Для окисления жирных кислот существует свой путь
Окисление жирных кислот (β-окисление)
Для преобразования энергии, заключенной в жирных кислотах, в энергию связей АТФ существует метаболический путь окисления жирных кислот до СО2 и воды, тесно связанный с циклом трикарбоновых кислот и дыхательной цепью. Этот путь называется β-окисление, т.к. происходит окисление 3-го углеродного атома жирной кислоты (β-положение) в карбоксильную группу, одновременно от кислоты отщепляется ацетильная группа, включающая С 1 и С 2 исходной жирной кислоты.
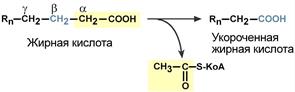
Элементарная схема β-окисления
Реакции β-окисления происходят в митохондриях большинства клеток организма (кроме нервных клеток). Для окисления используются жирные кислоты, поступающие в цитозоль из крови или появляющиеся при липолизе собственных внутриклеточных ТАГ. Суммарное уравнение окисления пальмитиновой кислоты выглядит следующим образом:
Пальмитоил-SКоА + 7ФАД + 7НАД + + 7Н2O + 7HS-KoA → 8Ацетил-SКоА + 7ФАДН2 + 7НАДН
Этапы окисления жирных кислот
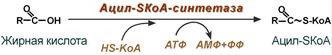
1. Прежде, чем проникнуть в матрикс митохондрий и окислиться, жирная кислота должна активироваться в цитозоле. Это осуществляется присоединением к ней коэнзима А с образованием ацил-SКоА. Ацил-SКоА является высокоэнергетическим соединением. Необратимость реакции достигается гидролизом дифосфата на две молекулы фосфорной кислоты.
Реакция активации жирной кислоты
2. Ацил-SКоА не способен проходить через митохондриальную мембрану, поэтому существует способ переноса жирной кислоты в комплексе с витаминоподобным веществом карнитином (витамин В11). На наружной мембране митохондрий имеется фермент карнитин-ацилтрансфераза I.
![]()
Карнитин-зависимый транспорт жирных кислот в митохондрию
3. После связывания с карнитином жирная кислота переносится через внутреннюю митохондриальную мембрану транслоказой . На внутренней стороне этой мембраны фермент карнитин-ацилтрансфераза II вновь образует ацил-SКоА, который вступает на путь β-окисления.
4. Процесс собственно β-окисления состоит из 4-х реакций, повторяющихся циклически. В них последовательно происходит окисление (ацил-SКоА-дегидрогеназа), гидратирование (еноил-SКоА-гидратаза) и вновь окисление 3-го атома углерода (гидроксиацил-SКоА-дегидрогеназа). В последней, трансферазной, реакции от жирной кислоты отщепляется ацетил-SКоА . К оставшейся (укороченной на два углерода) жирной кислоте присоединяется HS-КоА, и она возвращается к первой реакции. Все повторяется до тех пор, пока в последнем цикле не образуются два ацетил-SКоА.
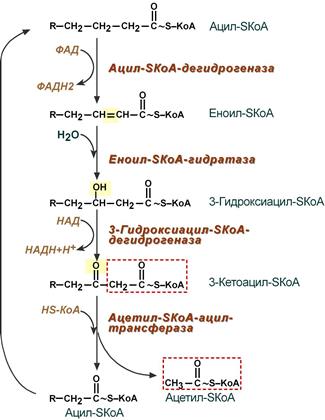
Последовательность реакций β-окисления жирных кислот
Расчет энергетического баланса β-окисления
Ранее при расчете эффективности окисления коэффициент P/O для НАДH принимался равным 3,0, для ФАДH2 – 2,0.
По современным данным значение коэффициента P/O для НАДH соответствует 2,5, для ФАДH2 – 1,5.
При расчете количества АТФ, образуемого при β-окислении жирных кислот необходимо учитывать:
- количество образуемого ацетил-SКоА – определяется обычным делением числа атомов углерода в жирной кислоте на 2.
- число циклов β-окисления . Число циклов β-окисления легко определить исходя из представления о жирной кислоте как о цепочке двухуглеродных звеньев. Число разрывов между звеньями соответствует числу циклов β-окисления. Эту же величину можно подсчитать по формуле (n/2 -1), где n – число атомов углерода в кислоте.
- число двойных связей в жирной кислоте. В первой реакции β-окисления происходит образование двойной связи при участии ФАД. Если двойная связь в жирной кислоте уже имеется, то необходимость в этой реакции отпадает и ФАДН2 не образуется. Количество недополученных ФАДН2 соответствует числу двойных связей. Остальные реакции цикла идут без изменений.
- количество энергии АТФ , потраченной на активацию (всегда соответствует двум макроэргическим связям).
Бета-окисление жирных кислот: стадии, реакции, продукты, регуляция
Бета-окисление жирных кислот: стадии, реакции, продукты, регуляция — Наука
Содержание:
В бета-окисление жирных кислот Это путь катаболизма (разложения) жирных кислот, основная функция которого — производство или «высвобождение» энергии, содержащейся в связях этих молекул.
Этот путь был открыт в 1904 году благодаря экспериментам, проведенным немцем Францем Кнопом, которые заключались в введении экспериментальным крысам жирных кислот, последняя метильная группа которых была модифицирована фенильной группой.
Кнуп ожидал, что продукты катаболизма этих «аналогов» жирных кислот пойдут путями, аналогичными путям окисления нормальных (немодифицированных природных) жирных кислот. Однако он обнаружил различия в получаемых продуктах в зависимости от числа атомов углерода жирных кислот.
Получив эти результаты, Кнуп предположил, что разложение происходит «ступенчато», начиная с «атаки» на β-углерод (тот, который находится в положении 3 относительно концевой карбоксильной группы), высвобождая фрагменты из двух атомов углерода.
Позже было показано, что для этого процесса требуется энергия в форме АТФ, которая вырабатывается в митохондриях, и что фрагменты из двух атомов углерода входят в цикл Кребса в виде ацетил-КоА.
Короче говоря, бета-окисление жирных кислот включает активацию концевой карбоксильной группы, перенос активированной жирной кислоты в митохондриальный матрикс и «ступенчатое» окисление два на два углерода карбоксильной группы.
Как и многие анаболические и катаболические процессы, этот путь регулируется, так как он заслуживает мобилизации «резервных» жирных кислот, когда другие катаболические пути недостаточны для удовлетворения энергетических потребностей клеток и тела.
Шаги и реакции
Жирные кислоты преимущественно находятся в цитозоле, независимо от того, поступают ли они из биосинтетических путей или из жировых отложений, которые накапливаются из принятой пищи (которая должна проникать в клетки).
— Активация жирных кислот и транспорт в митохондрии
Активация жирных кислот требует использования молекулы АТФ и связана с образованием конъюгатов ацилтиоэфира с коферментом А.
Эта активация катализируется группой ферментов, называемых ацетил-КоА-лигазами, специфичными для длины цепи каждой жирной кислоты. Некоторые из этих ферментов активируют жирные кислоты, поскольку они транспортируются в матрикс митохондрий, поскольку они встроены во внешнюю мембрану митохондрий.
Процесс активации происходит в два этапа: сначала производится ациладенилат из активированной жирной кислоты с помощью АТФ, где высвобождается молекула пирофосфата (PPi). Карбоксильная группа, активированная АТФ, затем подвергается атаке тиольной группы кофермента А с образованием ацил-КоА.
Транслокация ацил-КоА через митохондриальную мембрану достигается транспортной системой, известной как карнитиновый челнок.
— Бета-окисление насыщенных жирных кислот с четным числом атомов углерода
Разложение жирных кислот — это циклический путь, так как за высвобождением каждого фрагмента из двух атомов углерода немедленно следует другой, пока не будет достигнута полная длина молекулы. В этом процессе участвуют следующие реакции:
— Гидратация двойной связи.
— Дегидрирование гидроксильной группы.
— Фрагментация за счет атаки молекулы ацетил-КоА на β-углерод.
Реакция 1: первое дегидрирование
Он состоит из образования двойной связи между α-углеродом и β-углеродом путем удаления двух атомов водорода. Он катализируется ферментом ацил-КоА-дегидрогеназой, который образует молекулу транс∆2-еноил-S-КоА и молекулу FAD + (кофактор).
Реакции 2 и 3: гидратация и дегидрирование.
Гидратация катализируется еноил-КоА-гидратазой, в то время как дегидрирование опосредуется 3-гидроксиацил-КоА-дегидрогеназой, и последняя реакция зависит от кофактора НАД +.
Гидратация транс-2-еноил-S-CoA приводит к образованию 3-гидроксиацил-CoA, дегидрирование которого дает молекулу 3-кетоацил-CoA и NADH + H.
FADH2 и NADH, образующиеся в первых трех реакциях бета-окисления, повторно окисляются через цепь переноса электронов, благодаря чему они участвуют в производстве ATP, 2 молекулы для каждого FADH2 и 3 молекулы для каждого NADH.
Реакция 4: фрагментация
Каждый цикл бета-окисления, который удаляет молекулу с двумя атомами углерода, заканчивается «тиолитической» фрагментацией кетоуглерода, который подвергается атаке коэнзима А на связи между α и β атомами углерода.
Эта реакция катализируется ферментом β-кетотиолазой или тиолазой, а ее продуктами являются молекула ацил-КоА (активированная жирная кислота с двумя атомами углерода меньше) и одна — ацетил-КоА.
— Бета-окисление насыщенных жирных кислот с нечетным числом атомов углерода
В жирных кислотах с нечетным числом атомов углерода (которых не так много) молекула последнего цикла разложения имеет 5 атомов углерода, поэтому при ее фрагментации образуется молекула ацетил-КоА (которая входит в цикл Krebs) и еще один пропионил-КоА.
Пропионил-КоА должен быть карбоксилирован (реакция зависит от АТФ и бикарбоната) ферментом пропионил-КоА-карбоксилазой, с помощью которого образуется соединение, известное как D-метилмалонил-КоА, которое должно быть эпимеризовано до его L-формы.
Затем соединение, полученное в результате эпимеризации, превращается в сукцинил-КоА под действием фермента L-метилмалонил-КоА мутазы, и эта молекула, так же как ацетил-КоА, входит в цикл лимонной кислоты.
— Бета-окисление ненасыщенных жирных кислот
Многие клеточные липиды имеют цепи ненасыщенных жирных кислот, то есть они имеют одну или несколько двойных связей между их атомами углерода.
Окисление этих жирных кислот немного отличается от окисления насыщенных жирных кислот, поскольку два дополнительных фермента, еноил-КоА-изомераза и 2,4-диеноил-КоА-редуктаза, отвечают за устранение этих ненасыщенных соединений, так что эти жирные кислоты может быть субстратом для фермента еноил-КоА-гидратазы.
Эноил-КоА-изомераза действует на мононенасыщенные жирные кислоты (только с одной ненасыщенностью), в то время как фермент 2,4-диеноил-КоА-редуктаза реагирует с полиненасыщенными жирными кислотами (с двумя или более ненасыщенными).
— Бета внемитохондриальное окисление
Бета-окисление жирных кислот может также происходить внутри других цитозольных органелл, таких как пероксисомы, например, с той разницей, что электроны, которые переносятся на FAD +, доставляются не в дыхательную цепь, а непосредственно в кислород.
В результате этой реакции образуется перекись водорода (кислород уменьшается), соединение, которое удаляется ферментом каталазы, специфичным для этих органелл.
Продукты бета-окисления
Окисление жирных кислот дает гораздо больше энергии, чем расщепление углеводов. Основным продуктом бета-окисления является ацетил-КоА, образующийся на каждом этапе циклического участка пути, однако другими продуктами являются:
— AMP, H + и пирофосфат (PPi), образующиеся при активации.
— FADH2 и NADH для каждого произведенного ацетил-КоА.
— Сукцинил-КоА, АДФ, Pi, для жирных кислот с нечетной цепью.
Если мы рассмотрим в качестве примера полное бета-окисление пальмитиновой кислоты (пальмитата), жирной кислоты с 16 атомами углерода, количество производимой энергии эквивалентно более или менее 129 молекулам АТФ, которые поступают из 7 витков, которые он должен завершить. цикл.
Регулирование
Регулирование бета-окисления жирных кислот в большинстве клеток зависит от доступности энергии, связанной не только с углеводами, но и с самими жирными кислотами.
Животные контролируют мобилизацию и, следовательно, расщепление жиров посредством гормональных стимулов, которые в то же время контролируются такими молекулами, как, например, цАМФ.
В печени, главном органе расщепления жира, концентрация малонил-КоА чрезвычайно важна для регуляции бета-окисления; это первый субстрат, участвующий в пути биосинтеза жирных кислот.
Когда малонил-КоА накапливается в больших количествах, он способствует биосинтезу жирных кислот и ингибирует митохондриальный переносчик или челнок ацил-карнитина. Когда его концентрация уменьшается, ингибирование прекращается и активируется бета-окисление.
Ссылки
- Мэтьюз, К., ван Холде, К., и Ахерн, К. (2000). Биохимия (3-е изд.). Сан-Франциско, Калифорния: Пирсон.
- Нельсон, Д. Л., и Кокс, М. М. (2009). Принципы биохимии Ленингера. Издания Омега (5-е изд.).
- Рон, Дж. Д. (1998). Биохимия. Берлингтон, Массачусетс: Нил Паттерсон Паблишерс.
- Шульц, Х. (1991). Бета-окисление жирных кислот. Biochimica et Biophysica Acta, 1081, 109–120.
- Шульц, Х. (1994). Регулирование окисления жирных кислот в сердце. Критический обзор, 165–171.
- Шульц, Х. и Кунау, В. (1987). Бета-окисление ненасыщенных жирных кислот: новый путь. TIBS, 403–406.
50 отличных фраз аниме (вдохновляющих и запоминающихся)
ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТ
Установлено, что окисление жирных кислот протекает в печени, почках, скелетных и сердечной мышцах, в жировой ткани. В мозговой ткани скорость окисления жирных кислот весьма незначительна; основным источником энергии в мозговой ткани служит глюкоза.
В 1904 г. Ф. Кнооп (F. Knoop) выдвинул гипотезу β-окисления жирных кислот на основании опытов по скармливанию собакам различных жирных кислот, в которых один атом водорода в концевой метильной группе (ω-углеродного атома) был замещен радикалом (С6Н5–).
Ф. Кнооп высказал предположение, что окисление молекулы жирной кислоты в тканях организма происходит в β-положении. В результате от молекулы жирной кислоты последовательно отщепляются двууглеродные фрагменты со стороны карбоксильной группы.
Жирные кислоты, входящие в состав естественных жиров животных и растений, имеют четное число углеродных атомов. Любая такая кислота, от которой отщепляется по паре углеродных атомов, в конце концов проходит через стадию масляной кислоты. После очередного β-окисления масляная кислота становится ацетоуксусной. Последняя затем гидроли-зуется до двух молекул уксусной кислоты. Теория β-окисления жирных кислот, предложенная Ф. Кноопом, в значительной мере послужила основой современных представлений о механизме окисления жирных кислот.
Доставка жирных кислот к месту их окисления – к митохондриям – происходит сложным путем: при участии альбумина осуществляется транспорт жирной кислоты в клетку; при участии специальных белков (fatty acid binding proteins, FABP) – транспорт в пределах цитозоля; при участии карнитина – транспорт жирной кислоты из цитозоля в митохондрии.
Процесс окисления жирных кислот складывается из следующих основных этапов.
Активация жирных кислот. Свободная жирная кислота независимо от длины углеводородной цепи является метаболически инертной и не может подвергаться никаким биохимическим превращениям, в том числе окислению, пока не будет активирована. Активация жирной кислоты протекает на наружной поверхности мембраны митохондрий при участии АТФ, коэнзима A (HS-KoA) и ионов Mg 2+ . Реакция катализируется ферментом ацил-КоА-синтетазой:
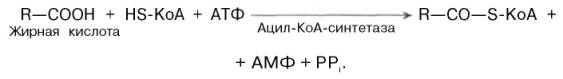
В результате реакции образуется ацил-КоА, являющийся активной формой жирной кислоты.
Считают, что активация жирной кислоты протекает в 2 этапа. Сначала жирная кислота реагирует с АТФ с образованием ациладенилата, представляющим собой эфир жирной кислоты и АМФ. Далее сульфгидрильная группа КоА действует на прочно связанный с ферментом ациладенилат с образованием ацил-КоА и АМФ.
Транспорт жирных кислот внутрь митохондрий. Коэнзимная форма жирной кислоты, в равной мере как и свободные жирные кислоты, не обладает способностью проникать внутрь митохондрий, где, собственно, и протекает их окисление. Переносчиком активированных жирных кислот
с длинной цепью через внутреннюю митохондриальную мембрану служит карнитин. Ацильная группа переносится с атома серы КоА на гидроксильную группу карнитина с образованием ацилкарнитина, который диффундирует через внутреннюю митохондриальную мембрану:
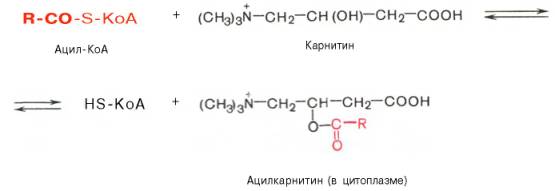
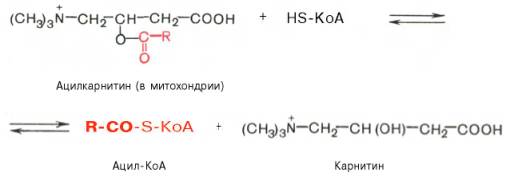
Реакция протекает при участии специфического цитоплазматического фермента карнитин-ацилтрансферазы. Уже на той стороне мембраны, которая обращена к матриксу, ацильная группа переносится обратно на КоА, что термодинамически выгодно, поскольку О-ацильная связь в кар-нитине обладает высоким потенциалом переноса группы. Иными словами, после прохождения ацилкарнитина через мембрану митохондрий происходит обратная реакция – расщепление ацилкарнитина при участии HS-KoA и митохондриальной карнитин-ацилтрансферазы:
Внутримитохондриальное окисление жирных кислот. Процесс окисления жирной кислоты в митохондриях клетки включает несколько последовательных энзиматических реакций.
Первая стадия дегидрирования. Ацил-КоА в митохондриях прежде всего подвергается ферментативному дегидрированию, при этом ацил-КоА теряет 2 атома водорода в α- и β-положениях, превращаясь в КоА-эфир ненасыщенной кислоты. Таким образом, первой реакцией в каждом цикле распада ацил-КоА является его окисление ацил-КоА-де-гидрогеназой, приводящее к образованию еноил-КоА с двойной связью между С-2 и С-3:
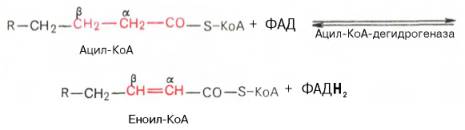
Существует несколько ФАД-содержащих ацил-КоА-дегидрогеназ, каждая из которых обладает специфичностью по отношению к ацил-КоА с определенной длиной углеродной цепи.
Стадия гидратации. Ненасыщенный ацил-КоА (еноил-КоА) при участии фермента еноил-КоА-гидратазы присоединяет молекулу воды. В результате образуется β-оксиацил-КоА (или 3-гидроксиацил-КоА):
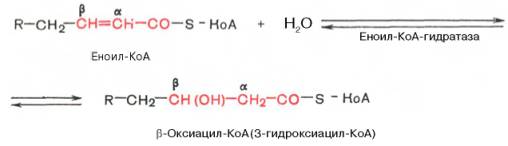
Заметим, что гидратация еноил-КоА стереоспецифична, подобно гидратации фумарата и аконитата (см. с. 348). В результате гидратации транс-Δ 2 -двойной связи образуется только L-изомер 3-гидроксиацил-КоА.
Вторая стадия дегидрирования. Образовавшийся β-оксиацил-КоА (3-гидроксиацил-КоА) затем дегидрируется. Эту реакцию катализируют НАД + -зависимые дегидрогеназы:
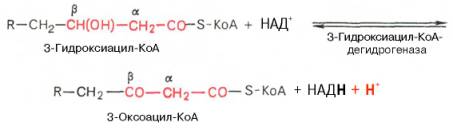
Тиолазная реакция. В ходе предыдущих реакций происходило окисление метиленовой группы при С-3 в оксогруппу. Тиолазная реакция представляет собой расщепление 3-оксоацил-КоА с помощью тиоловой группы второй молекулы КоА. В результате образуется укороченный на два углеродных атома ацил-КоА и двууглеродный фрагмент в виде ацетил-КоА. Данная реакция катализируется ацетил-КоА-ацилтрансферазой (β-ке-тотиолазой):
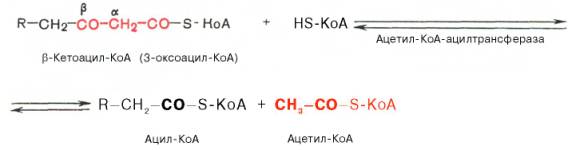
Образовавшийся ацетил-КоА подвергается окислению в цикле трикар-боновых кислот, а ацил-КоА, укоротившийся на два углеродных атома, снова многократно проходит весь путь β-окисления вплоть до образования бутирил-КоА (4-углеродное соединение), который в свою очередь окисляется до 2 молекул ацетил-КоА (рис. 11.2). Например, при окислении пальмитиновой кислоты (С16) повторяется 7 циклов β-окисления. Запомним, что при окислении жирной кислоты, содержащей п углеродных
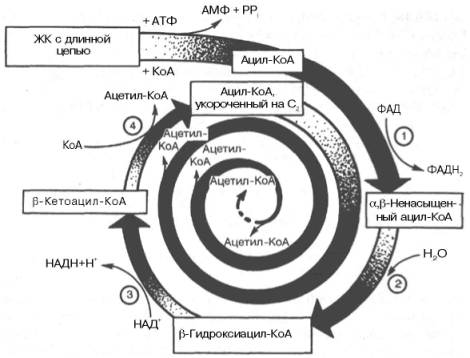
Рис. 11.2. Окисление жирной кислоты («спираль Линена»). Подробно представлен первый цикл окисления — укорочение цепи жирной кислоты на два углеродных атома. Остальные циклы аналогичны первому (по А.Н. Климову и Н.Г. Никульче-вой).
1 — ацил-КоА-дегидрогеназа (КФ 1.3.99.3) ; 2 — еноил-КоА-гидратаза (КФ 4.2.1.17.); 3 — β-гидро-ксиацил-КоА-дегидрогеназа (КФ 1.1.1.35); 4 — тиолаза (КФ 2.3.1.9).
атомов, происходит n/2–1 цикл β-окисления (т.е. на один цикл меньше, чем n/2, так как при окислении бутирил-КоА сразу происходит образование 2 молекул ацетил-КоА) и всего получится п/2 молекул ацетил-КоА. Следовательно, суммарное уравнение β-окисления активированной кислоты можно записать так:
Пальмитоил-КоА + 7ФАД + 7НАД + + 7Н2O + 7HS-KoA –>
–> 8Ацетил-КоА + 7ФАДН2 + 7НАДН + 7Н + .
Баланс энергии. При каждом цикле β-окисления образуются одна молекула ФАДН2 и одна молекула НАДН. Последние в процессе окисления в дыхательной цепи и сопряженного с ним фосфорилирования дают: ФАДН2 – 2 молекулы АТФ и НАДН – 3 молекулы АТФ, т.е. в сумме за один цикл образуется 5 молекул АТФ. При окислении пальмитиновой кислоты образуется 5 х 7 = 35 молекул АТФ. В процессе β-окисления пальмитиновой кислоты образуется 8 молекул ацетил-КоА, каждая из которых, «сгорая» в цикле трикарбоновых кислот, дает 12 молекул АТФ, а 8 молекул ацетил-КоА дадут 12 х 8 = 96 молекул АТФ.
Таким образом, всего при полном β-окислении пальмитиновой кислоты образуется 35 + 96 = 131 молекула АТФ. С учетом одной молекулы АТФ, потраченной в самом начале на образование активной формы пальмитиновой кислоты (пальмитоил-КоА), общий энергетический выход при полном окислении одной молекулы пальмитиновой кислоты в условиях животного организма составит 131 – 1 = 130 молекул АТФ. Изменение свободной энергии ΔF при полном сгорании 1 моля пальмитиновой кислоты составляет 2338 ккал, а богатая энергией фосфатная связь АТФ характеризуется величиной 7,6 ккал/моль. Нетрудно подсчитать, что примерно 990 ккал (7,6 х 130), или 42% от всей потенциальной энергии пальмитиновой кислоты при ее окислении в организме, используется для ресинтеза АТФ, а оставшаяся часть, очевидно, теряется в виде тепла.
Следовательно, эффективность накопления энергии в результате окисления жирных кислот при стандартных условиях составляет
http://ru1.warbletoncouncil.org/beta-oxidacion-de-acidos-grasos-13572
http://xumuk.ru/biologhim/159.html