Фотосинтез
Типы питания
По типу питания живые организмы делятся на автотрофы, гетеротрофы и миксотрофы. Автотрофы (греч. αὐτός — сам + τροφ — пища) — организмы, которые самостоятельно способны синтезировать органические вещества из неорганических. Гетеротрофы (греч. ἕτερος — иной + τροφή — пища) — организмы, использующие для питания готовые органические вещества.
Наконец, миксотрофы (греч. μῖξις — смешение + τροφή — пища) — организмы, которые могут использовать как гетеротрофный, так и автотрофный способ питания. К примеру, эвглена зеленая на свету начинает фотосинтезировать, а в темноте питается гетеротрофно.

Фотосинтез
Фотосинтез (греч. φῶς — свет и σύνθεσις — синтез) — сложный химический процесс преобразования энергии квантов света в энергию химических связей. В результате фотосинтеза происходит синтез органических веществ из неорганических.

Этот процесс уникален и происходит только в растительных клетках, а также у некоторых бактерий. Фотосинтез осуществляется при участии хлорофилла (греч. χλωρός — зелёный и φύλλον — лист) — зеленого пигмента, окрашивающего органы растений в зеленый цвет. Существуют и другие вспомогательные пигменты, которые вместе с хлорофиллом выполняют светособирающую или светозащитную функции.
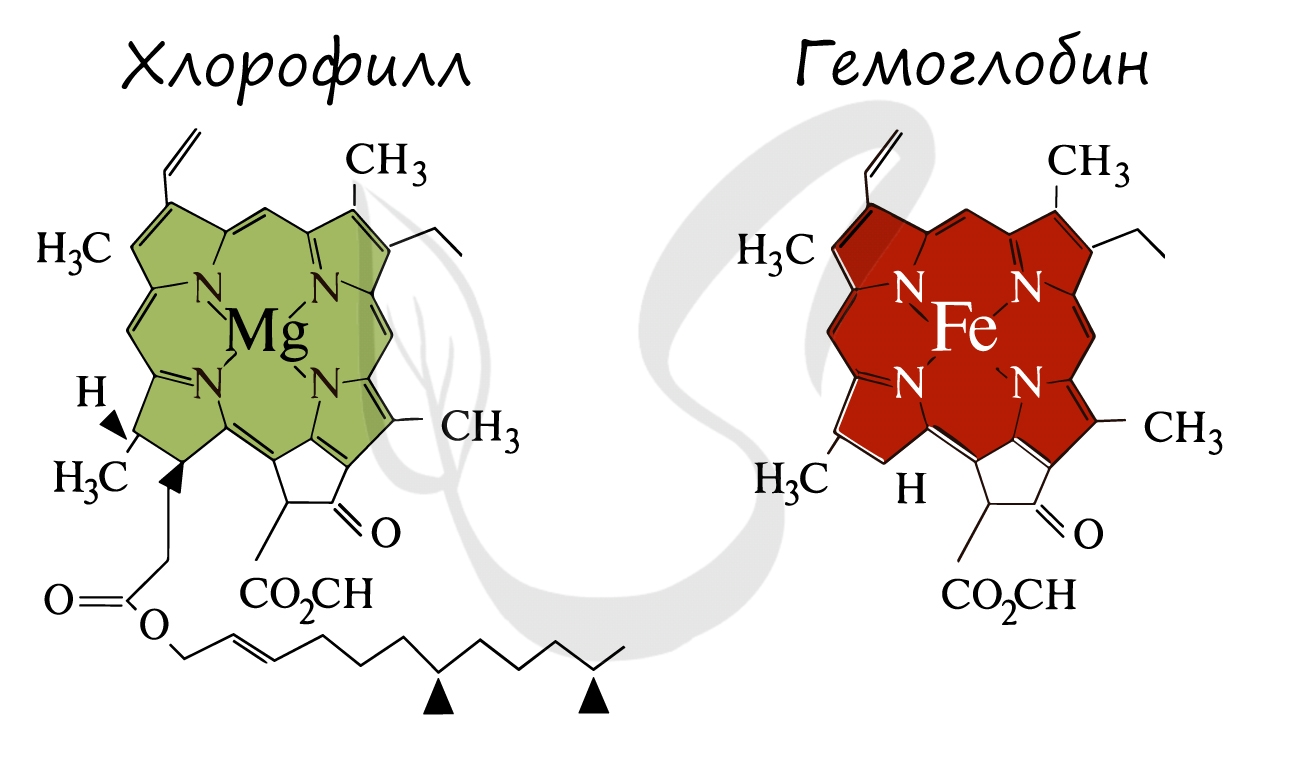
Ниже вы увидите сравнение строения хлорофилла и гемоглобина. Обратите внимание, что в центре молекулы хлорофилла находится ион Mg.
В высшей степени гениально значение процесса фотосинтеза подчеркнул русский ученый К.А. Тимирязев: «Все органические вещества, как бы они ни были разнообразны, где бы они ни встречались, в растении ли, в животном или человеке, прошли через лист, произошли от веществ, выработанных листом. Вне листа или, вернее, вне хлорофиллового зерна в природе не существует лаборатории, где бы выделялось органическое вещество. Во всех других органах и организмах оно превращается, преобразуется, только здесь оно образуется вновь из вещества неорганического»
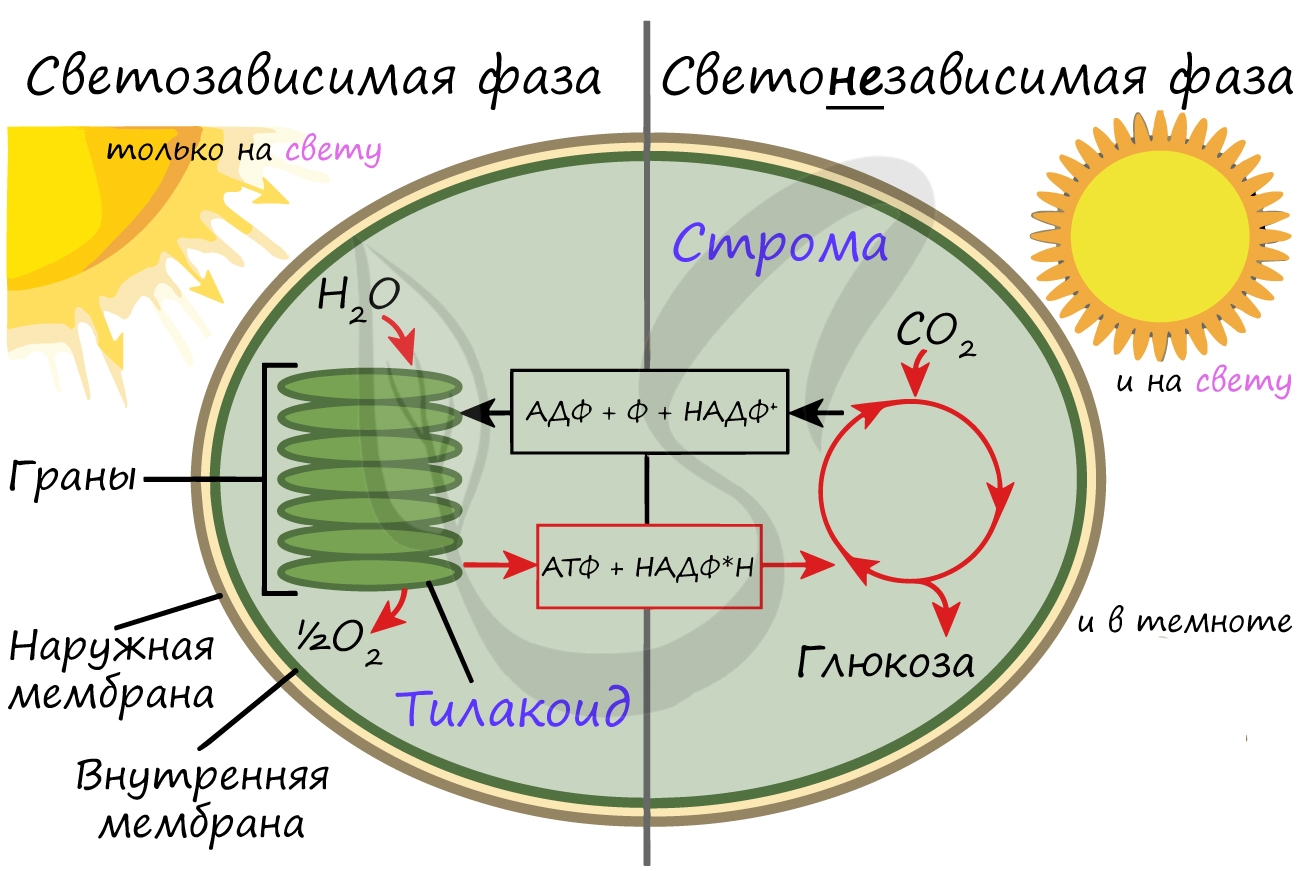
Более подробно мы обсудим значение фотосинтеза в завершение этой статьи. Фотосинтез состоит из двух фаз: светозависимой (световой) и светонезависимой (темновой). Я рекомендую использовать названия светозависимая и светонезависимая, так как они способствуют более глубокому (и правильному!) пониманию фотосинтеза.
Светозависимая фаза (световая)
Эта фаза происходит только на свету на мембранах тилакоидов в хлоропластах. В ней принимают участие различные ферменты, белки-переносчики, молекулы АТФ-синтетазы и зеленый пигмент хлорофилл.
Хлорофилл выполняет две функции: поглощения и передачи энергии. При воздействии кванта света хлорофилл теряет электрон, переходя в возбужденное состояние. С помощью переносчиков электроны скапливаются с наружной поверхности мембраны тилакоидов, тем временем внутри тилакоида происходит фотолиз воды (разложение под действием света):
Гидроксид-ионы отдают лишний электрон, превращаясь в реакционно способные радикалы OH, которые собираются вместе и образуют молекулу воды и свободный кислород (это побочный продукт, который в дальнейшем удаляется в ходе газообмена).
Образовавшиеся при фотолизе воды протоны (H + ) скапливаются с внутренней стороны мембраны тилакоидов, а электроны — с внешней. В результате по обе стороны мембраны накапливаются противоположные заряды.
При достижении критической разницы, часть протонов проталкивается на внешнюю сторону мембраны через канал АТФ-синтетазы. В результате этого выделяется энергия, которая может быть использована для фосфорилирования молекул АДФ:
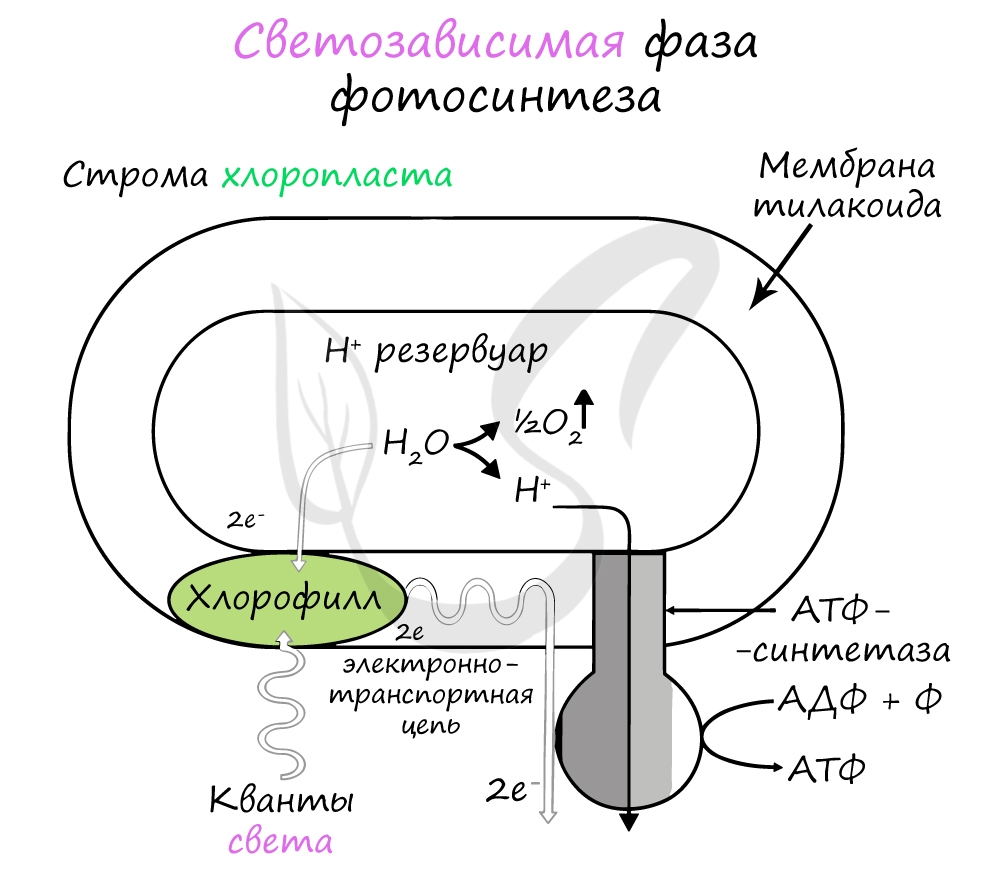
Протоны, попав на поверхность мембраны тилакоидов, соединяются с электронами и образуют атомарный водород, который используется для восстановления молекулы-переносчика НАДФ (никотинамиддинуклеотидфосфат). Благодаря этому окисленная форма — НАФД + превращается в восстановленную — НАДФ∗H2.
Предлагаю создать квинтэссенцию из полученных нами знаний. Итак, в результате светозависимой фазы фотосинтеза образуются:
- Свободный кислород O2 — в результате фотолиза воды
- АТФ — универсальный источник энергии
- НАДФ∗H2 — форма запасания атомов водорода
Кислород удаляется из клетки как побочный продукт фотосинтеза, он совершенно не нужен растению. АТФ и НАДФ∗H2 в дальнейшем оказываются более полезны: они транспортируются в строму хлоропласта и принимают участие в светонезависимой фазе фотосинтеза.
Светонезависимая (темновая) фаза
Светонезависимая фаза происходит в строме (матриксе) хлоропласта постоянно: и днем, и ночью — вне зависимости от освещения.
При участии АТФ и НАДФ∗H2 происходит восстановление CO2 до глюкозы C6H12O6. В светонезависимой фазе происходит цикл Кальвина, в ходе которого и образуется глюкоза. Для образования одной молекулы глюкозы требуется 6 молекул CO2, 12 НАДФ∗H2 и 18 АТФ.
Таким образом, в результате темновой (светонезависимой) фазы фотосинтеза образуется глюкоза, которая в дальнейшем может быть преобразована в крахмал, служащий для запасания питательных веществ у растений.
Значение фотосинтеза
Значение фотосинтеза невозможно переоценить. Уверенно утверждаю: именно благодаря этому процессу жизнь на Земле приобрела такие чудесные и изумительные формы, какие мы видим вокруг себя: удивительные растения, прекрасные цветы и самые разнообразные животные.
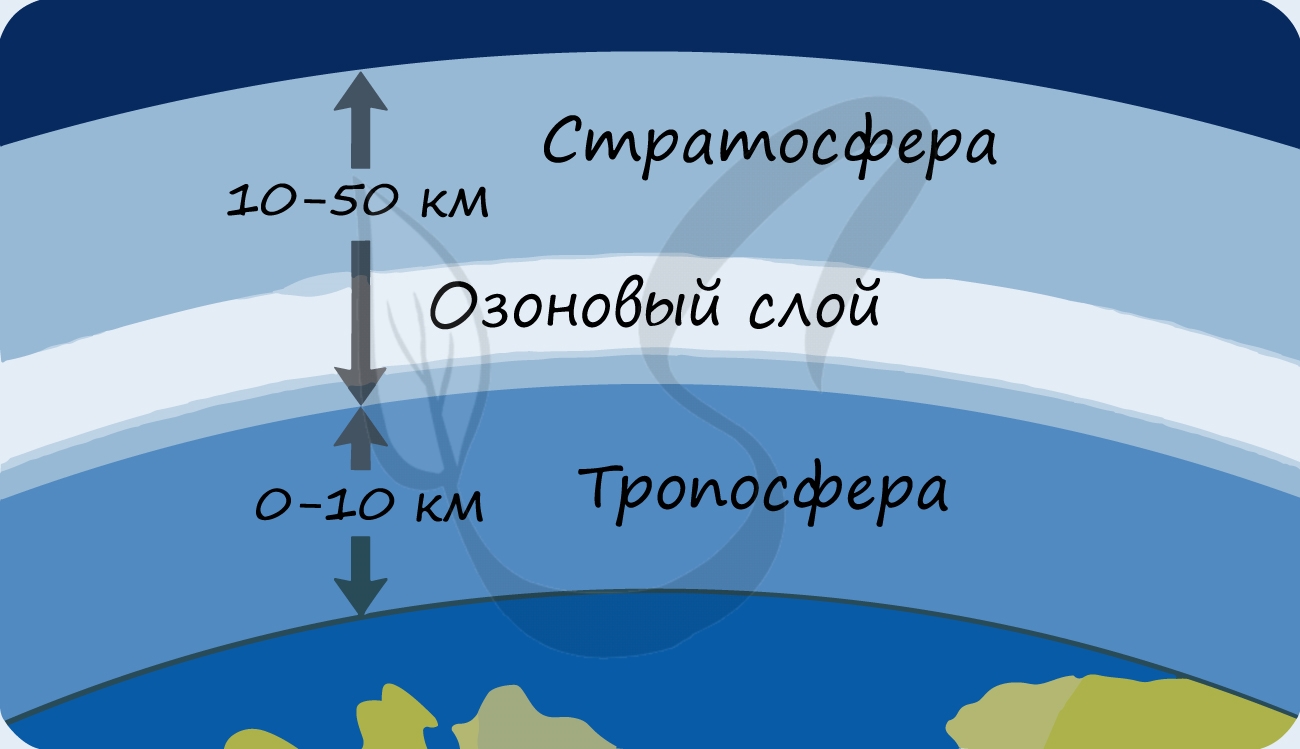
В разделе эволюции мы уже обсуждали, что изначально в составе атмосферы Земли не было кислорода: миллиарды лет назад его начали вырабатывать первые фотосинтезирующие бактерии — сине-зеленые водоросли (цианобактерии). Постепенно кислород накапливался, и со временем на Земле стало возможно аэробное (кислородное) дыхание. Возник озоновый слой, защищающий все живое на нашей планете от губительного ультрафиолета.
Говоря о роли фотосинтеза, выделим следующие функции, объединяющиеся в так называемую космическую роль растений. Итак, растения за счет фотосинтеза:
- Синтезируют органические вещества, являющиеся пищей для всего живого на планете
- Преобразуют энергию света в энергию химических связей, создают органическую массу
- Растения поддерживают определенный процент содержания O2 в атмосфере, очищают ее от избытка CO2
- Способствуют образованию защитного озонового экрана, поглощающего губительное для жизни ультрафиолетовое излучение

Хемосинтез (греч. chemeia – химия + synthesis — синтез)
Хемосинтез — автотрофный тип питания, который характерен для некоторых микроорганизмов, способных создавать органические вещества из неорганических. Это осуществляется за счет энергии, получаемой при окислении других неорганических соединений (железо- , азото-, серосодержащих веществ).
Хемосинтез был открыт русским микробиологом С.Н. Виноградским в 1888 году. Большинство хемосинтезирующих бактерий относится к аэробам, для жизни им необходим кислород.
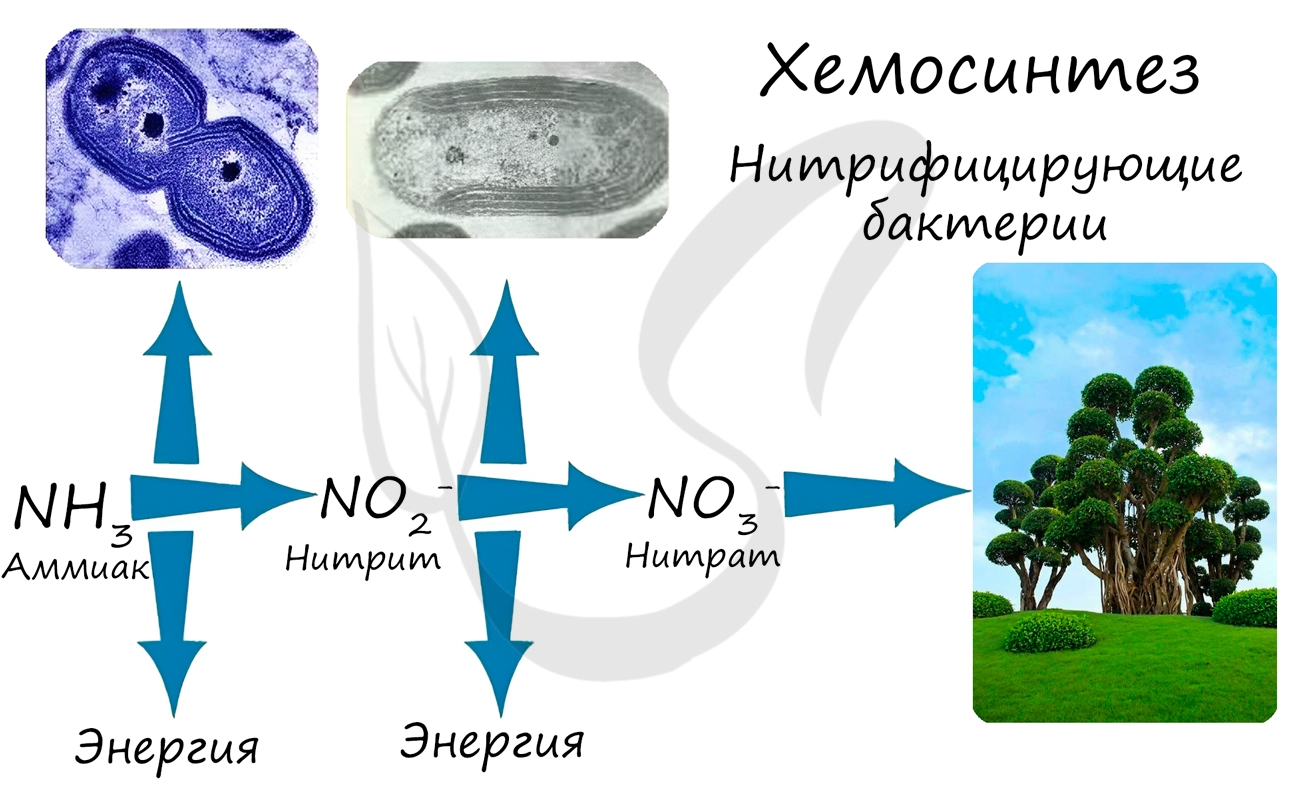
При окислении неорганических веществ выделяется энергия, которую организмы запасают в виде энергии химических связей. Так нитрифицирующие бактерии последовательно окисляют аммиак до нитрита, а затем — нитрата. Нитраты могут быть усвоены растениями и служат удобрением.
Помимо нитрифицирующих бактерий, встречаются:
- Серобактерии — окисляют H2S —> S 0 —> (S +4 O3) 2- —> (S +6 O4) 2-
- Железобактерии — окисляют Fe +2 —>Fe +3
- Водородные бактерии — окисляют H2 —> H +1 2O
- Карбоксидобактерии — окисляют CO до CO2
Значение хемосинтеза
Хемосинтезирующие бактерии являются неотъемлемым звеном круговорота в природе таких элементов как: азот, сера, железо.
Нитрифицирующие бактерии обеспечивают переработку (нейтрализацию) ядовитого вещества — аммиака. Они также обогащают почву нитратами, которые очень важны для нормального роста и развития растений.
Усвоение нитратов происходит за счет клубеньковых бактерий на корнях бобовых растений, однако важно помнить, что клубеньковые (азотфиксирующие) бактерии, в отличие от нитрифицирующих бактерий, питаются гетеротрофно.
© Беллевич Юрий Сергеевич 2018-2022
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Световая стадия фотосинтеза
Основой жизнедеятельности растений является фотосинтез. В процессе фотосинтеза в тканях растения из неорганических веществ образуются органические. Как и любая химическая реакция синтеза, фотосинтез проходит с поглощением энергии.
Что такое световая фаза фотосинтеза
Фотосинтез – сложный процесс, состоящий из 2 фаз: световой и темновой. Реакции протекающие в световой фазе фотосинтеза могут проходить только при освещении. Для темновой фазы свет не важен, она может проходить в любое время. Ниже представлена схема световой и темновой фазы фотосинтеза.
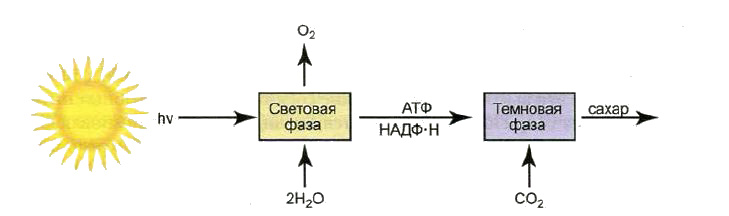
В течение световой стадии, растение захватывает фотоны света с помощью специальных светособирающих комплексов. Энергия фотонов необходима для прохождения процесса распада воды на кислород и водород.
Этот процесс, который происходит в световую фазу фотосинтеза называется фотолизом воды. Далее происходит образование конечных продуктов световой фазы, которые необходимы для прохождения реакций темновой фазы. Световая энергия накапливается в виде АТФ – аденозинтрифосфата.
Водород после фотолиза воды соединяется с ферментом НАДФ (никотинамидадениндинуклеотидфосфат), образуя НАДФН, который является источником водорода для дальнейших химических реакций.
Кислород при фотолизе воды выделяется в атмосферу. Таким образом, результатом световой фазы фотосинтеза является: распад воды под воздействием световой энергии с образованием конечных продуктов АТФ и НАДФН, использующихся для синтеза органики и свободного кислорода.
Где происходит световая фаза
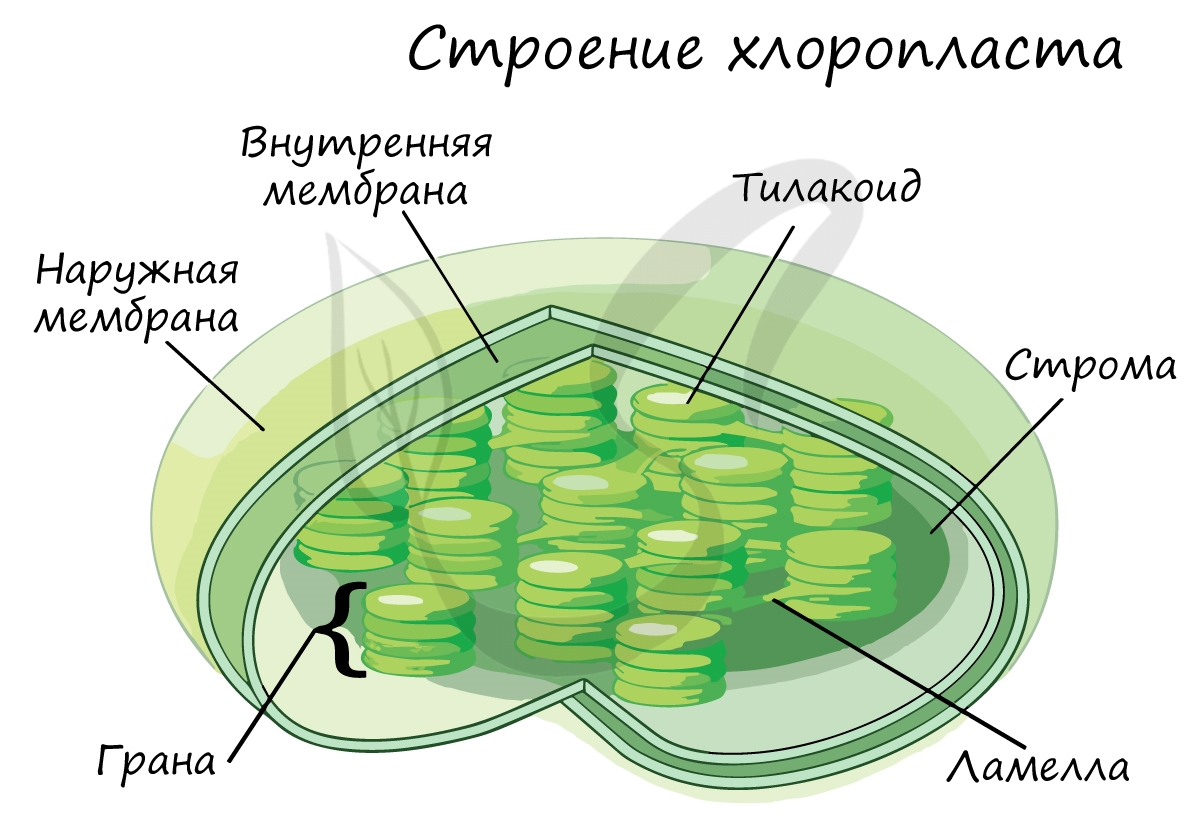
Все процессы происходящие в световой стадии и темновой фазе фотосинтеза протекают в специальных клеточных структурах, называемых хлоропластами. Хлоропласт – зеленая пластида, внутри которой содержится хлорофилл. В растительной клетке содержится большое количество хлоропластов, необходимых для прохождения химических реакций фотосинтеза.
Различные фазы процесса проходят в разных частях хлоропласта. Эта пластида имеет сложную структуру, в ее состав входит большое количество тилакоидов. Тилакоиды – особые структуры внутри хлоропласта, отвечающие за преобразование световой энергии. Тилакоиды, расположенные рядом, образуют стопки – граны.
Световая фаза фотосинтеза происходит в гранах тилакоидов, на их мембранах и во внутритилакоидном пространстве. В этом особенность световой фазы фотосинтеза и этим она отличается от темновой, во время которой химические реакции протекают в строме хлоропласта – плотном веществе между тилакоидами.
Фотохимическая суть процесса
Основным процессом световой фазы фотосинтеза является фотолиз воды, представленный следующим уравнением:
Молекулы хлорофилла имеют свойство возбуждаться, и терять электроны при попадании на них кванта света. Эти электроны оседают на внешней стороне мембран, заряжая их отрицательно.
Сами молекулы хлорофилла, потерявшие электрон, восстанавливаются, отбирая электроны у воды, находящейся внутри тилакоида. При фотолизе вода расщепляется на следующие компоненты:
- Протоны водорода;
- Электроны водорода;
- Кислород.
Кислород является побочным продуктом фотосинтеза и не участвует в дальнейших процессах. Он выводится наружу из ткани растения и поступает в атмосферу. Протоны накапливаются в специальном протонном резервуаре, находящемся внутри тилакоида.
Протоны заряжают внутреннюю часть тилакоидной мембраны положительно. Таким образом, мембраны тилакоидов имеют отрицательный заряд с внешней стороны, а положительный – с внутренней. Эти заряды постепенно увеличиваются по мере накопления протонов и электронов.
Разность потенциалов между внешней и внутренней поверхностью мембраны должна достигать не менее 200mВ, чтобы начался процесс образования конечных продуктов световой стадии. Только в этом случае протоны начнут проходить сквозь каналы АТФ-синтазы, находящиеся в тилакоидных мембранах.
2Н + + 2е — + НАДФ → НАДФН
Светособирающие комплексы
Только небольшая часть молекул хлорофилла поглощает энергию, отдавая электроны. Они находятся в реакционных центрах и называются молекулами-ловушками. Остальная же часть этого пигмента собрана в светособирающие комплексы, задачей которых является не поглощение, а передача энергии.
Для чего нужны светособирающие комплексы? Если бы каждая молекула хлорофилла улавливала свет, то такая работа была бы крайне неэффективной. Процесс возбуждения и потери электрона проходил бы очень редко, а структура передачи электронов была бы слишком сложной из-за очень большого количества молекул.
На самом деле существует очень мало молекул, поглощающих энергию и отдающих электроны. На каждую из них приходится до 300 молекул, собранных в светособирающие комплексы по антенному типу. Они расположены на нескольких уровнях.
Конечно, происходит передача не квантов света, а только энергии, полученной при поглощении света. Таким образом, хлорофилл может не только поглощать световую энергию, но и передавать ее.
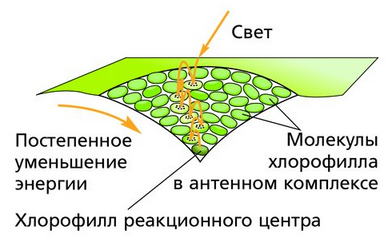
На самом нижнем уровне светособирающего комплекса находится 1 молекула-ловушка. Энергия поступает к ней со всего антенного комплекса. Передача энергии происходит с определенными потерями ее количества.
Но молекула-ловушка получает энергию в десятки или даже в сотни раз чаще, чем молекулы, расположенные на самом высоком уровне. Молекулы-ловушки входят в состав фотосистем, которые участвуют в транспорте электронов во время световой стадии фотосинтеза.
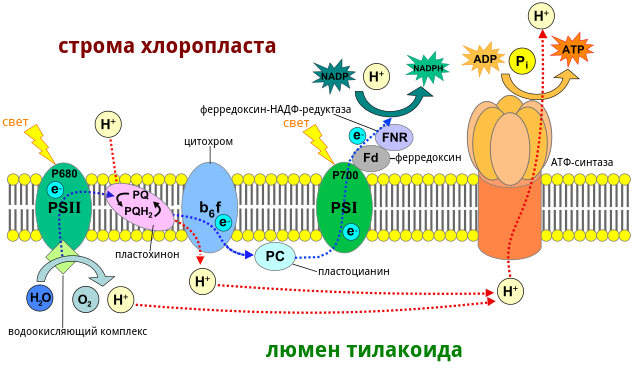
Основные компоненты цепи переноса электронов
В течение световой стадии происходит перенос электронов от тилакоидных фотосистем с помощью промежуточных веществ-переносчиков до образования конечных продуктов фазы. Электрон-транспортная цепь имеет сложную структуру и множество компонентов.
Основными компонентами цепи переноса электронов являются:
- Фотосистема 1;
- Фотосистема 2;
- Комплекс цитохромов b6-f;
- Вещества-переносчики;
- НАДФ-редуктаза.
Фотосистема 1 способна поглощать свет с длиной волны 700 нм, фотосистема 2 – 680 нм. Фотосистемы работают параллельно. При поглощении света фотосистемы отдают электроны на вещества-переносчики или акцепторы. В электрон-транспортной цепи задействовано множество акцепторов, которые захватывают электроны и отдают их другому компоненту цепи.
Когда фотосистема 2 теряет электрон под воздействием света, он сначала попадает на акцептор феофитин. Далее в его транспорте принимает участие целый ряд акцепторов, последним из которых является пластоцианин. Далее электрон попадает в фотосистему 1, восполняя электрон, отданный этой фотосистемой под воздействием кванта света.
Фотосистема 1 отдает электрон на акцептор ферредоксин. Отсюда он поступает в последний компонент цепи НАДФ-редуктазу. Здесь образуется в световую фазу фотосинтеза вещество НАДФН. Недостачу электронов фотосистема 1 восполняет за счет электронов, приходящих от фотосистемы 2.
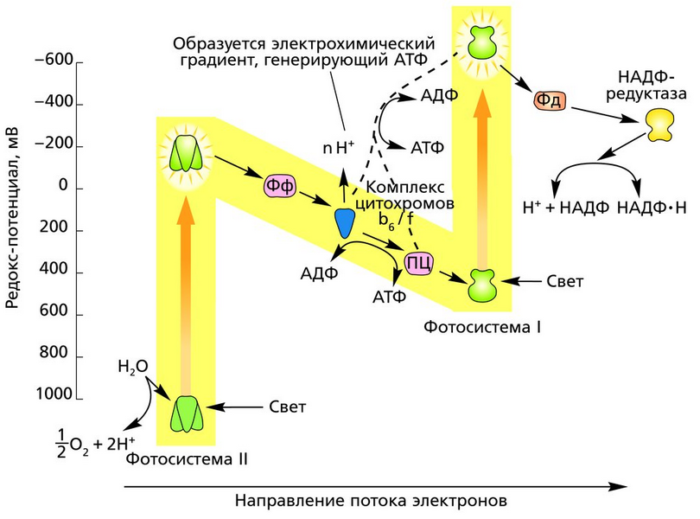
Особое значение в электрон-транспортной цепи имеет комплекс цитохромов b6-f. Электроны, проходя через этот комплекс, многократно взаимодействуют с акцептором пластохиноном. При этом комплекс цитохромов увеличивает количество, не только электронов, но также и протонов, что повышает эффективность световой стадии.
Продукты световой стадии
При прохождении этапа световой фазы фотосинтеза образуются следующие продукты, необходимые для синтеза органики в дальнейших темновых реакциях: АТФ и НАДФН. АТФ – источник биохимической энергии. Эта молекула синтезируется из АДФ при поглощении энергии движущегося протона.
Формулу синтеза АТФ во время световой фазы фотосинтеза можно представить в следующем формуле:
АДФ + ортофосфорная кислота + энергия → АТФ + Н2О
Синтезированный АТФ может участвовать во всех химических реакциях, для прохождения которых необходима энергия. При взаимодействии с водой происходит обратная реакция с выделением энергии.
АТФ вновь расщепляется на АДФ и ортофосфорную кислоту:
АТФ + H2O → АДФ + ортофосфорная кислота + энергия
Для образования органических веществ при фотосинтезе такая энергетическая составляющая крайне необходима, так как синтез органики требует поглощения большого количества энергии. НАДФН – восстановленный фермент, который является источником водорода. Он используется в химических процессах темновой фазы, где отдает водород и превращается в фермент НАДФ.
Фотосинтез
Фотосинтез — это преобразование энергии света в энергию химических связей органических соединений.
Фотосинтез характерен для растений, в том числе всех водорослей, ряда прокариот, в том числе цианобактерий, некоторых одноклеточных эукариот.
В большинстве случаев при фотосинтезе в качестве побочного продукта образуется кислород (O2). Однако это не всегда так, поскольку существует несколько разных путей фотосинтеза. В случае выделения кислорода его источником является вода, от которой на нужды фотосинтеза отщепляются атомы водорода.
Фотосинтез состоит из множества реакций, в которых участвуют различные пигменты, ферменты, коферменты и др. Основными пигментами являются хлорофиллы, кроме них — каротиноиды и фикобилины.
В природе распространены два пути фотосинтеза растений: C3 и С4. У других организмов есть своя специфика реакций. Все, что объединяет эти разные процессы под термином «фотосинтез», – во всех них в общей сложности происходит преобразование энергии фотонов в химическую связь. Для сравнения: при хемосинтезе происходит преобразование энергии химической связи одних соединений (неорганических) в другие — органические.
Выделяют две фазы фотосинтеза — световую и темновую. Первая зависит от светового излучения (hν), которое необходимо для протекания реакций. Темновая фаза является светонезависимой.
У растений фотосинтез протекает в хлоропластах. В результате всех реакций образуются первичные органические вещества, из которых потом синтезируются углеводы, аминокислоты, жирные кислоты и др. Обычно суммарную реакцию фотосинтеза пишут в отношении глюкозы — наиболее распространенного продукта фотосинтеза:
Атомы кислорода, входящие в молекулу O2, берутся не из углекислого газа, а из воды. Углекислый газ – источник углерода, что более важно. Благодаря его связыванию у растений появляется возможность синтеза органики.
Представленная выше химическая реакция есть обобщенная и суммарная. Она далека от сути процесса. Так глюкоза не образуется из шести отдельных молекул углекислоты. Связывание CO2 происходит по одной молекуле, которая сначала присоединяется к уже существующему пятиуглеродному сахару.
Для прокариот характерны свои особенности фотосинтеза. Так у бактерий главный пигмент — бактериохлорофилл, и не выделяется кислород, так как водород берется не из воды, а часто из сероводорода или других веществ. У сине-зеленых водорослей основным пигментом является хлорофилл, и при фотосинтезе выделяется кислород.
Световая фаза фотосинтеза
В световой фазе фотосинтеза происходит синтез АТФ и НАДФ·H2 за счет лучистой энергии. Это происходит на тилакоидах хлоропластов, где пигменты и ферменты образуют сложные комплексы для функционирования электрохимических цепей, по которым передаются электроны и отчасти протоны водорода.
Электроны в конечном итоге оказываются у кофермента НАДФ, который, заряжаясь отрицательно, притягивает к себе часть протонов и превращается в НАДФ·H2. Также накопление протонов по одну сторону тилакоидной мембраны и электронов по другую создает электрохимический градиент, потенциал которого используется ферментом АТФ-синтетазой для синтеза АТФ из АДФ и фосфорной кислоты.
Главными пигментами фотосинтеза являются различные хлорофиллы. Их молекулы улавливают излучение определенных, отчасти разных спектров света. При этом некоторые электроны молекул хлорофилла переходят на более высокий энергетический уровень. Это неустойчивое состояние, и по-идее электроны путем того же излучения должны отдать в пространство полученную из вне энергию и вернуться на прежний уровень. Однако в фотосинтезирующих клетках возбужденные электроны захватываются акцепторами и с постепенным уменьшением своей энергии передаются по цепи переносчиков.
На мембранах тилакоидов существуют два типа фотосистем, испускающих электроны при действия света. Фотосистемы представляют собой сложный комплекс большей частью хлорофильных пигментов с реакционным центром, от которого и отрываются электроны. В фотосистеме солнечный свет ловит множество молекул, но вся энергия собирается в реакционном центре.
Электроны фотосистемы I, пройдя по цепи переносчиков, восстанавливают НАДФ.
Энергия электронов, оторвавшихся от фотосистемы II, используется для синтеза АТФ. А сами электроны фотосистемы II заполняют электронные дырки фотосистемы I.
Дырки второй фотосистемы заполняются электронами, образующимися в результате фотолиза воды. Фотолиз также происходит при участии света и заключается в разложении H2O на протоны, электроны и кислород. Именно в результате фотолиза воды образуется свободный кислород. Протоны участвуют в создании электрохимического градиента и восстановлении НАДФ. Электроны получает хлорофилл фотосистемы II.
Примерное суммарное уравнение световой фазы фотосинтеза:
H2O + НАДФ + 2АДФ + 2Ф → ½O2 + НАДФ · H2 + 2АТФ
Циклический транспорт электронов
Выше описана так называемый нецикличная световая фаза фотосинтеза. Есть еще циклический транспорт электронов, когда восстановления НАДФ не происходит. При этом электроны от фотосистемы I уходят на цепь переносчиков, где идет синтез АТФ. То есть эта электрон-транспортная цепь получает электроны из фотосистемы I, а не II. Первая фотосистема как бы реализует цикл: в нее возвращаются ей же испускаемые электроны. По дороге они тратят часть своей энергии на синтез АТФ.
Фотофосфорилирование и окислительное фосфорилирование
Световую фазу фотосинтеза можно сравнить с этапом клеточного дыхания — окислительным фосфорилированием, которое протекает на кристах митохондрий. Там тоже происходит синтез АТФ за счет передачи электронов и протонов по цепи переносчиков. Однако в случае фотосинтеза энергия запасается в АТФ не для нужд клетки, а в основном для потребностей темновой фазы фотосинтеза. И если при дыхании первоначальным источником энергии служат органические вещества, то при фотосинтезе – солнечный свет. Синтез АТФ при фотосинтезе называется фотофосфорилированием, а не окислительным фосфорилированием.
Темновая фаза фотосинтеза
Впервые темновую фазу фотосинтеза подробно изучили Кальвин, Бенсон, Бэссем. Открытый ими цикл реакций в последствии был назван циклом Кальвина, или C3-фотосинтезом. У определенных групп растений наблюдается видоизмененный путь фотосинтеза – C4, также называемый циклом Хэтча-Слэка.
В темновых реакциях фотосинтеза происходит фиксация CO2. Темновая фаза протекает в строме хлоропласта.
Восстановление CO2 происходит за счет энергии АТФ и восстановительной силы НАДФ·H2, образующихся в световых реакциях. Без них фиксации углерода не происходит. Поэтому хотя темновая фаза напрямую не зависит от света, но обычно также протекает на свету.
Цикл Кальвина
Первая реакция темновой фазы – присоединение CO2 (карбоксилирование) к 1,5-рибулезобифосфату (рибулезо-1,5-дифосфат) – РиБФ. Последний представляет собой дважды фосфорилированную рибозу. Данную реакцию катализирует фермент рибулезо-1,5-дифосфаткарбоксилаза, также называемый рубиско.
В результате карбоксилирования образуется неустойчивое шестиуглеродное соединение, которое в результате гидролиза распадается на две трехуглеродные молекулы фосфоглицериновой кислоты (ФГК) – первый продукт фотосинтеза. ФГК также называют фосфоглицератом.
ФГК содержит три атома углерода, один из которых входит в состав кислотной карбоксильной группы (-COOH):
Из ФГК образуется трехуглеродный сахар (глицеральдегидфосфат) триозофосфат (ТФ), включающий уже альдегидную группу (-CHO):
ФГК (3-кислота) → ТФ (3-сахар)
На данную реакцию затрачивается энергия АТФ и восстановительная сила НАДФ · H2. ТФ — первый углевод фотосинтеза.
После этого большая часть триозофосфата затрачивается на регенерацию рибулозобифосфата (РиБФ), который снова используется для связывания CO2. Регенерация включает в себя ряд идущих с затратой АТФ реакций, в которых участвуют сахарофосфаты с количеством атомов углерода от 3 до 7.
В таком круговороте РиБФ и заключается цикл Кальвина.
Из цикла Кальвина выходит меньшая часть образовавшегося в нем ТФ. В перерасчете на 6 связанных молекул углекислого газа выход составляет 2 молекулы триозофосфата. Суммарная реакция цикла с входными и выходными продуктами:
При этом в связывании участвую 6 молекул РиБФ и образуется 12 молекул ФГК, которые превращаются в 12 ТФ, из которых 10 молекул остаются в цикле и преобразуются в 6 молекул РиБФ. Поскольку ТФ — это трехуглеродный сахар, а РиБФ — пятиуглеродный, то в отношении атомов углерода имеем: 10 * 3 = 6 * 5. Количество атомов углерода, обеспечивающих цикл не изменяется, весь необходимый РиБФ регенерируется. А шесть вошедших в цикл молекул углекислоты затрачиваются на образование двух выходящих из цикла молекул триозофосфата.
На цикл Кальвина в расчете на 6 связанных молекул CO2 затрачивается 18 молекул АТФ и 12 молекул НАДФ · H2, которые были синтезированы в реакциях световой фазы фотосинтеза.
Расчет ведется на две выходящие из цикла молекулы триозофосфата, так как образующаяся в последствии молекула глюкозы, включает 6 атомов углерода.
Триозофосфат (ТФ) — конечный продукт цикла Кальвина, но его сложно назвать конечным продуктом фотосинтеза, так как он почти не накапливается, а, вступая в реакции с другими веществами, превращается в глюкозу, сахарозу, крахмал, жиры, жирные кислоты, аминокислоты. Кроме ТФ важную роль играет ФГК. Однако подобные реакции происходят не только у фотосинтезирующих организмов. В этом смысле темновая фаза фотосинтеза – это то же самое, что цикл Кальвина.
Из ФГК путем ступенчатого ферментативного катализа образуется шестиуглеродный сахар фруктозо-6-фосфат, который превращается в глюкозу. В растениях глюкоза может полимеризоваться в крахмал и целлюлозу. Синтез углеводов похож на процесс обратный гликолизу.
Фотодыхание
Кислород подавляет фотосинтез. Чем больше O2 в окружающей среде, тем менее эффективен процесс связывания CO2. Дело в том, что фермент рибулозобифосфат-карбоксилаза (рубиско) может реагировать не только с углекислым газом, но и кислородом. В этом случае темновые реакции несколько иные.
Содержащая пять атомов углерода молекула рибулозобифосфата реагирует уже не с CO2, а с O2. В результате чего образуются по одной молекуле фосфогликолата (C2) и фосфоглицериновой кислоты (C3), а не две ФГК как обычно.
Фосфогликолат — это фосфогликолевая кислота. От нее сразу отщепляется фосфатная группа, и она превращается в гликолевую кислоту (гликолат). Для его «утилизации» снова нужен кислород. Поэтому чем больше в атмосфере кислорода, тем больше он будет стимулировать фотодыхание и тем больше растению будет требоваться кислорода, чтобы избавиться от продуктов реакции.
Фотодыхание — это зависимое от света потребление кислорода и выделение углекислого газа. То есть обмен газов происходит как при дыхании, но протекает в хлоропластах и зависит от светового излучения. От света фотодыхание зависит лишь потому, что рибулозобифосфат образуется только при фотосинтезе.
При фотодыхании происходит возврат атомов углерода из гликолата в цикл Кальвина в виде фосфоглицериновой кислоты (фосфоглицерата).
2 Гликолат (С2) → 2 Глиоксилат (С2) →2 Глицин (C2) — CO2 → Серин (C3) →Гидроксипируват (C3) → Глицерат (C3) → ФГК (C3)
Как видно, возврат происходит не полный, так как один атом углерода теряется при превращении двух молекул глицина в одну молекулу аминокислоты серина, при этом выделяется углекислый газ.
Кислород необходим на стадиях превращения гликолата в глиоксилат и глицина в серин.
Превращения гликолата в глиоксилат, а затем в глицин происходят в пероксисомах, синтез серина в митохондриях. Серин снова поступает в пероксисомы, где из него сначала получается гидрооксипируват, а затем глицерат. Глицерат уже поступает в хлоропласты, где из него синтезируется ФГК.
Фотодыхание характерно в основном для растений с C3-типом фотосинтеза. Его можно считать вредным, так как энергия бесполезно тратится на превращения гликолата в ФГК. Видимо фотодыхание возникло из-за того, что древние растения были не готовы к большому количеству кислорода в атмосфере. Изначально их эволюция шла в атмосфере богатой углекислым газом, и именно он в основном захватывал реакционный центр фермента рубиско.
C4-фотосинтез, или цикл Хэтча-Слэка
Если при C3-фотосинтезе первым продуктом темновой фазы является фосфоглицериновая кислота, включающая три атома углерода, то при C4-пути первыми продуктами являются кислоты, содержащие четыре атома углерода: яблочная, щавелевоуксусная, аспарагиновая.
С4-фотосинтез наблюдается у многих тропических растений, например, сахарного тростника, кукурузы.
С4-растения эффективнее поглощают оксид углерода, у них почти не выражено фотодыхание.
Растения, в которых темновая фаза фотосинтеза протекает по C4-пути, имеют особое строение листа. В нем проводящие пучки окружены двойным слоем клеток. Внутренний слой — обкладка проводящего пучка. Наружный слой — клетки мезофилла. Хлоропласты клеток слоев отличаются друг от друга.
Для мезофильных хлоропласт характерны крупные граны, высокая активность фотосистем, отсутствие фермента РиБФ-карбоксилазы (рубиско) и крахмала. То есть хлоропласты этих клеток адаптированы преимущественно для световой фазы фотосинтеза.
В хлоропластах клеток проводящего пучка граны почти не развиты, зато высока концентрация РиБФ-карбоксилазы. Эти хлоропласты адаптированы для темновой фазы фотосинтеза.
Углекислый газ сначала попадает в клетки мезофилла, связывается с органическими кислотами, в таком виде транспортируется в клетки обкладки, освобождается и далее связывается также, как у C3-растений. То есть C4-путь дополняет, а не заменяет C3.
В мезофилле CO2 присоединяется к фосфоенолпирувату (ФЕП) с образованием оксалоацетата (кислота), включающего четыре атома углерода:
Реакция происходит при участии фермента ФЕП-карбоксилазы, обладающего более высоким сродством к CO2, чем рубиско. К тому же ФЕП-карбоксилаза не взаимодействует с кислородом, а значит не затрачивается на фотодыхание. Таким образом, преимущество C4-фотосинтеза заключается в более эффективной фиксации углекислоты, увеличению ее концентрации в клетках обкладки и следовательно более эффективной работе РиБФ-карбоксилазы, которая почти не расходуется на фотодыхание.
Оксалоацетат превращается в 4-х углеродную дикарбоновую кислоту (малат или аспартат), которая транспортируется в хлоропласты клеток обкладки проводящих пучков. Здесь кислота декарбоксилируется (отнятие CO2), окисляется (отнятие водорода) и превращается в пируват. Водород восстанавливает НАДФ. Пируват возвращается в мезофилл, где из него регенерируется ФЕП с затратой АТФ.
Оторванный CO2 в хлоропластах клеток обкладки уходит на обычный C3-путь темновой фазы фотосинтеза, т. е. в цикл Кальвина.
Фотосинтез по пути Хэтча-Слэка требует больше энергозатрат.
Считается, что C4-путь возник в эволюции позже C3 и во многом является приспособлением против фотодыхания.
http://florist-club.com/fotosintez/svetovaya-faza.html
http://biology.su/molecular/photosynthesis