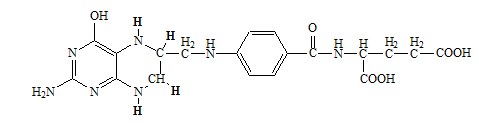
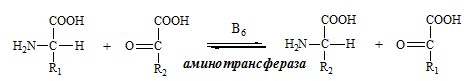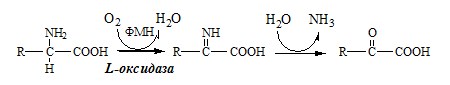Синтез глицина


Синтез аминокислоты глицина постоянно происходит в организме человека, обеспечивая расходным материалом для производства белков, гормонов, биологически-активных молекул. Прочитайте статью до конца, и вы узнаете механизмы реакций, протекающих в организме для синтеза глицина. Из чего образуется глицин, где он образуется и какие дополнительные компоненты необходимы для синтеза глицина. С вами Галина Батуро и аминокислота глицин.
Синтез глицина

Аминокислота глицин является заменимой протеиногенной аминокислотой. Это значит, что глицин входит в состав белков. Он присутствует в больших количествах в желатине, в виде амида он является составной частью гормонов вазопрессина и окситоцина. Вазопрессин, как явствует из его названия, это гормон, сужающий сосуды и поднимающий артериальное давление. Окситоцин – это гормон, способствующий сокращению гладкой мускулатуры, в большом количестве он выделяется во время родов, заставляя сокращаться матку и выталкивать плод из чрева.
Глицин входит в состав глутатиона, гиппуровой и гликохолевой кислот. В организме человека также вырабатывается N-метил-производное глицина – саркозин. Это производное холина и аминокислоты метионина.
Глицин является источником таких важных метаболитов, как креатин, пуриновые основания и порфирины, из которых образуется белок крови гемоглобин.
Будучи заменимой аминокислотой, глицин может легко синтезироваться в организме. Он образуется в процессе следующих реакций: расщепление аминокислоты серина, синтез из воды и аммиака, аминирование глиоксиловой кислоты, деметилирование саркозина.
Синтез глицина из серина
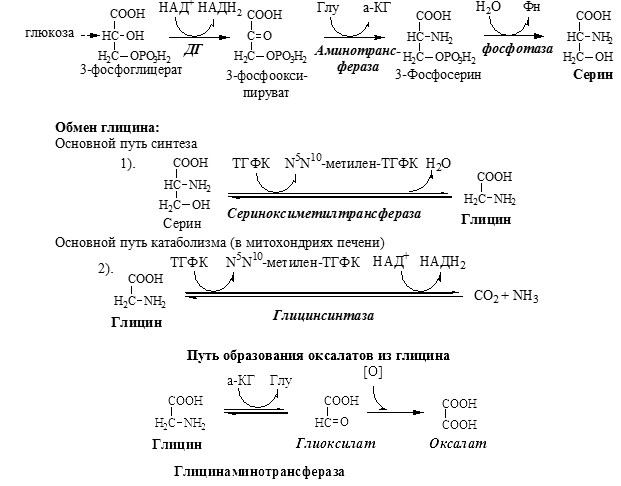
90% глицина синтезируется в организме из аминокислоты серин, тоже заменимой и протеиногенной. Углеродный скелет серин получает от 3-фосфоглицерата, промежуточного продукта распада глюкозы, а аминную голову предоставляет глутаминовая кислота. Казалось бы, при таких делах организм не должен бы испытывать недостаток глицина: глюкозу мы всегда получаем в избытке, и глутаминовая кислота, которая глутамат, обильно представлена в рационе. Однако засада подстерегает, где не ждешь. И называется она витамин B9, иначе говоря, фолиевая кислота, которую по идее мы должны получать со свежими листьями и травками. В нашей полосе, где девять месяцев зима, это особенно актуально.
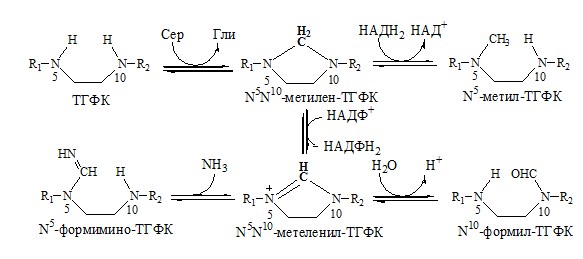
В синтезе глицина задействована активная форма витамина B9 (фолиевой кислоты) – Н4-Фолат, он же ТетраГидроФолиевая Кислота (ТГФК).

Образование ТГФК из фолиевой кислоты происходит в печени, это сложный каскад реакций, в которых задействованы особые ферменты, коферментом которых выступает НАДФ. Образовавшаяся ТГФК вступает в реакцию с серином при участии фермента СеринОксиМетилТрансфераза.

ТГФК принимает на себя метильную группу CH3, находящуюся в β-положении, и превращается N 5 N 10 МетиленН4Фолат, а спиртовая группа отщепляется в виде воды. Что значит это зубодробительное название? Фолат – это сокращенно фолиевая кислота, ибо соединение имеет гидроксильный хвост COOH. Метилен – означает, что соединение приняло на себя метильную группу CH2, причем приняли ее молекулы азота N в положении 5 и 10.

Реакция легко обратима, т.е. глицин может стать источником серина. В этом случае N 5 N 10 МетиленН4Фолат отдает метильную группу глицину, а вода станет источником спиртовой группы для серина. N 5 N 10 МетиленН4Фолат, образующийся вместе с глицином, быстренько превращается в N 5 МетилН4Фолат, который задействован в обезвреживании страшного гомоцистеина, превращая его в нужную аминокислоту метионин. В реакции принимает участие активная форма витамина B12 метилкобаламин.

Синтез глицина из треонина
Долгое время считалось, что распад треонина с образованием глицина идет в клетках печени (гепатоцитах) под воздействием фермента ТреонинАльдолазы. В учебниках рисовали красивое уравнение реакции, приведенное ниже. Не обманывайтесь, насчет обратимости реакции. Уже тогда подчеркивалось, что реакция в живых организмах преимущественно идет в сторону распада треонина с образованием глицина и ацетальдегида. Обратный синтез треонина в живых организмах не наблюдался.

В настоящее время стало известно, что фермент ТреонинАльдолаза расщепляет с образованием глицина не L-треонин, содержащийся в белках, а стереоизомер алло-треонин, который в синтезе белков не участвует (1).
Образование глицина при распаде треонина долгое время считался возможным в митохондриях под воздействием фермента ТреонинДегидрогеназы, активность которого зависит от НАД. В результате образуется аминоацетон, который окисляется до α-АминоАцетоУксусной Кислоты, а та, в свою очередь является предшественницей глицина. Следует отметить, что человек в процессе эволюции утратил способность к синтезу ТреонинДегидрогеназы, следовательно образование глицина из треонина в человеческом организме не возможно (2).
В печени человека в процессе обезвреживания глиоксиловой кислоты (о чем дальше), треонин может переаминироваться под воздействием фермента КинуренинАминоТрансферазы, который оказался идентичным СеринПируватАминоТрансферазе и АланинГлиоксилАминоТрансферазе. В пероксисомах печени в присутствии ПиридоксальФосфата (активная форма витамина В 6) треонин соединяется с глиоксиловой кислотой, в результате чего образуется глицин и α-Кето-β-АминоМасляная кислота.

Синтез глицина из углекислого газа и аммиака
В печени позвоночных (и человека) при участии фермента ГлицинСинтазы глицин может образовываться из углекислого газа, аммиака, а также активной формы витамина В9 — N 5 N 10 МетиленН4Фолат (см. реакцию 1) и НАДН+Н. Реакция обратима, т.е. глицин может деградировать до углекислого газа и аммиака с образованием метилированной формы Фолата и НАДН+Н. Реакция идет в присутствии активной формы витамина B6 пиридоксальфосфата. Таким образом, для синтеза глицина нужно два витамина: фолиевая кислота (B9) и пиридоксин (B6). Фолиевая кислота содержится в свежей зелени, а также в печени, но вот беда, при тепловой обработке она разрушается. Пиридоксин содержится в семенах подсолнечника, отрубном хлебе, фасоли, красной морской рыбе и других продуктах.

Синтез глицина из глиоксиловой кислоты
Глиоксиловая кислота — это жуткий яд, угнетающий тканевое дыхание. В больших количествах она содержится в незрелых фруктах, именно поэтому их не следует употреблять в пищу, особенно при проблемах с печенью и поджелудочной. В 30-е годы, когда в Среднем Поволжье разразился голод, мой двоюродный дед, будучи подростком, умер, поев незрелых яблок. Незрелые яблоки оказались соблазнительной пищей, с которой истощенный организм не справился. В другой раз чуть не отправился в кроличий рай мой домашний питомец, который дорвался до незрелых яблок, а я не сразу сообразила, что это не самая подходящая для него пища. Откачав крола, и вспомнив печальную историю двоюродного деда, я крепко-накрепко уяснила, что незрелые яблоки есть ни в коем случае нельзя. Теперь я знаю, почему – из-за высокого содержания глиоксиловой кислоты.
Глиоксиловая кислота также образуется в процессе биотрансформации этиленгликоля – яда, который добавляют в антифризы – жидкости-незамерзайки. При случайном (а иногда и не случайном) попадании внутрь, печень пытается обезвредить этиленгликоль, но в результате получаются соединения еще более ядовитые, и одним из них является глиоксиловая кислота.
В небольших количествах глиоксиловая кислота образуется, как побочный продукт, на пути образования холина из серина. Холин нам нужен, ибо из него получается нейромедиатор ацетилхолин.
Фермент декарбоксилаза откусывает у серина карбоксильный хвост, в результате чего получается аминоспирт 2-аминоэтанол и выделяется углекислый газ.

2-аминоэтанол может пойти на синтез холина и далее на синтез нейромедиатора ацетилхолина, а может превратиться в гликолевый альдегид, лишившись аминной головы в ходе окислительного дезаминирования.

Гликолевый альдегид – ядовитое вещество, которое надо немедленно обезвредить. Образуется он не только из 2-аминоэтанола на пути превращения серина, но также при распаде пуриновых оснований (каркаса молекул ДНК и РНК – генетических матриц клеток) и при альтернативном пути гликолиза – распаде сахаров с выходом энергии. Таким образом, имеется 3 источника естественного образования гликолевого альдегида:
- превращение аминокислоты серин с образованием 2-аминоэтанола, который дает гликолевый альдегид
- распад пуринов: ксантин преобразуется в соль мочевой кислоты, которая декарбоксилируется, т.е. теряет карбоновый хвост, превращаясь в аллантоин и аллантоиновую кислоты, а те гидролизируются до мочевины и гликолевого альдегида
- альтернативный гликолиз: глюкоза превращается во фруктозу-1,6-бисфосфат, а ту фермент кетолаза превращает в гликолевый альдегид.
Гликолевый альдегид образуется также при переработке этиленгликоля в печени, при отравлении этим соединением, причем парадоксально, продукты метаболизма опаснее самого яда.
Гликолевый альдегид окисляется ферментом АльдегидОксидазой до гликолевой кислоты, а та, в свою очередь окисляется ферментом ЛактатДегидрогеназой до глиоксиловой кислоты.

Все эти вещества являются ядами, угнетающими тканевое дыхание и синтез белка, они блокируют митохондриальный транспорт электронов, разобщают окисление и фосфорилирование, т.е. химическая энергия, выделяющаяся при сгорании органических молекул, рассеивается в виде тепла, а не используется на работу биохимического конвейера.
Организм очень хитро придумал, преобразовывать яды в полезное вещество глицин. Этим занимаются печеночные клетки в особых органеллах – микротельцах (пероксисомах).

Основная реакция обезвреживания глиоксиловой кислоты – это соединение с аланином.
Донором аминной группы в этой реакции выступает аминокислота аланин, которая превращается в пируват. Реакция идет при участии фермента АланинГлиоксилатАминоТрансферазы в сопровождении активной формы витамина В6 – ПиридоксальФосфаста.

Глиоксиловая кислота и орнитин
Другой реакцией обезвреживания глиоксиловой кислоты является соединение с орнитином, и на выходе получается глицин и γ-полуальдегид глутамиловой кислоты. Реакция активно идет в печени.

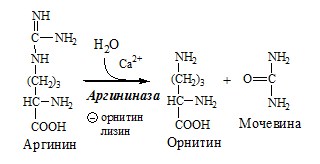
Где взять орнитин? Орнинин образуется из аргинина в процессе обезвреживания мочевины.
Прямой синтез глицина из глиоксиловой кислоты
Глицин может образовываться прямым синтезом из глиоксиловой кислоты. В этом случае аминную голову он берет у всевездесущего глутамата, который переходит в α-кетоглутарат. Реакция идет при участии фермента ГлицинАминоТрансферазы. Обратите внимание, что реакция обратима, т.е. глицин вполне может выступать источником глиоксиловой кислоты, и это не есть айс.

Глицин, таким образом, синтезируется, как конечный метаболит на пути обезвреживания гликолевого альдегида, гликолевой кислоты и глиоксиловой кислоты. Коль речь зашла об отравлении, надо сказать, что глиоксиловая кислота может превращаться в щавелевую кислоту, а та, поймав кальций, выпадает в осадок, образуя оксалаты – кальциевые соли щавелевой кислоты. Оксалаты представляют собой игольчатые кристаллы, они травмируют мочевыводящие протоки, образуют почечные камни. Образование большого количества оксалатов приводит к нарушению функции почек при отравлении этиленгликолем.
К чему это я? А к тому, что в некоторых случаях глицин выступает источником глиоксиловой кислоты, со всеми последствиями в виде образования в почках осксалатных камней.
Синтез глицина из саркозина
Саркозин – важный участник биохимического конвейера, в котором он задействован, как донор одноуглеродной группы в реакциях транметилирования, т.е. обмена метильными остатками CH3. Саркозин образуется в процессе распада холина. Превращения саркозина тесно связаны с глицином. Саркозин образуется из глицина, как непосредственно, так и через длинную биохимическую цепочку, в начале которой глицин превращается в серин, дальше через ряд соединений образуется холин, а из того при распаде получается саркозин.
У млекопитающихся (и человека) обнаружена размещенная в митохондриях ферментная система, которая разлагает саркозин с образованием глицина. 90% саркозина разлагается в митохондриях печени, а 10% — в почках. Реакция идет под действием фермента СаркозинДегидрогеназы, причем фермент работает при содействии апофермента, связанного с мембраной митохондрий. Его активность проявляется только в присутствии специфического флавопротеида ФАД (активной формы витамина В2 – рибофлавина), необходимого для переноса электронов. Эта реакция может идти как в присутствии, так и без ТетраГидроФолата (ТГФК – активной формы витамина В9 – фолиевой кислоты). В анаэробных условиях, т.е. без кислорода, ТГФК в реакции не участвует, в результате образуется свободный формальдегид, ядовитое вещество, которое срочно требуется обезвредить. В присутствии кислорода в реакцию вступает ТГФК, который забирает углеродный остаток СН3 у формальдегида, превращаясь в уже известный N 5 N 10 МетиленН4Фолат с выделением воды.
Точный механизм реакции не известен. Общая схема выглядит следующим образом:

Реакция в бескислородной среде протекает в два этапа. При участии кислорода и ТГФК – в три.
Первый этап: перенос водорода с N-метильной группы саркозина на ФАД, что позволяет воде атаковать образовавшийся положительно-заряженный карбокатион, с образованием промежуточного соединения. Происходит отсечение метильной головы, иначе говоря, деметилирование N-метильной группы на саркозине. Восстановленный на первой стадии ФАД Н — окисляется кислородом с образованием перекиси водорода.

Промежуточное соединение без ТГФК окисляется в глицин с выходом формальдегида.

На третьем этапе происходит обезвреживание формальдегида, для чего нужна ТГФК (активная форма фолиевой кислоты – витамина В9). ТГФК принимает на себя одноуглеродный остаток от формальдегида и превращается в N 5 ,N 10 -МетиленТетраГидроФолат.
Превращение происходит в четыре этапа с выделением воды.


Заключение
Глицин – активный участник биохимического конвейера, задействованный в переносе одноуглеродного остатка CH3, т.е. в реакциях метилирования. Обмен глицина тесно связан с обменом другой аминокислоты – серина и активной формы витамина B9 – фолиевой кислоты ТГФК. Биологический смысл данных процессов в передаче одноуглеродного остатка по биохимическому конвейеру, при этом глицин выступает источником одноуглеродного остатка, а ТГФК – передающим звеном, при этом обезвреживается образующийся в процессе биосинтеза опасный гомоцистеин с образованием безопасной аминокислоты метионин.
Глицин является конечным безопасным продуктом при обезвреживании клеточных ядов, имеющих как биосинтетическое происхождение, т.е. образующихся внутри организма при работе биохимического конвейера, так и пищевое происхождение, т.е. поступающих в организм извне, с пищей. Это гликолевый альдегид, гликолевая кислота и глиоксиловая кислота. В результате преобразований, происходящих в печени, клеточные яды превращаются в безопасный глицин.
Глицин также является конечным безопасным продуктом при утилизации холина и саркозина, в процессе которой образуются клеточный яд – формальдегид.
Распад биологически-активных молекул приводит к образованию опасных для организма веществ, требующих обезвреживания. Организм затрачивает усилия в виде энергии и работы ферментов и витаминов для того, чтобы запустить вторичное использование отработанных молекул, превращая яды в глицин. Глицин в свою очередь связывает обмен аминокислот, пуриновых оснований, порфиринов и, через аминокислоту серин, подпитывает углеводный обмен, давая организму энергию.
- Стр. 6. Малиновский А.В. «Переаминирование треонина»
- Стр. 15. Малиновский А.В. «Переаминирование треонина»
_» style=’position: relative; display: inline-block; /* height: 24px; */ /* padding: 0 12px 0 27px; */ /* border-radius: 12px; */ cursor: pointer; /* background: #21A5D8; */ text-decoration: none; padding: 2px 8px 2px 29px; /* font-size: 14px; */ /* font-family: Arial,sans-serif; */ /* color: #FFF; */ line-height: 25px; margin: 6px; color: #000; background: #fff; border: 1px solid #ccc; border-radius: 3px; vertical-align: middle; font-family: «Helvetica Neue»,Arial,sans-serif; font-size: 13px; line-height: 20px; cursor: pointer; line-height: 19px;’>
Acetyl
Наведите курсор на ячейку элемента, чтобы получить его краткое описание.
Чтобы получить подробное описание элемента, кликните по его названию.
| H + | Li + | K + | Na + | NH4 + | Ba 2+ | Ca 2+ | Mg 2+ | Sr 2+ | Al 3+ | Cr 3+ | Fe 2+ | Fe 3+ | Ni 2+ | Co 2+ | Mn 2+ | Zn 2+ | Ag + | Hg 2+ | Pb 2+ | Sn 2+ | Cu 2+ | |
| OH — | Р | Р | Р | Р | Р | М | Н | М | Н | Н | Н | Н | Н | Н | Н | Н | — | — | Н | Н | Н | |
| F — | Р | М | Р | Р | Р | М | Н | Н | М | М | Н | Н | Н | Р | Р | Р | Р | Р | — | Н | Р | Р |
| Cl — | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Н | Р | М | Р | Р |
| Br — | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Н | М | М | Р | Р |
| I — | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | ? | Р | ? | Р | Р | Р | Р | Н | Н | Н | М | ? |
| S 2- | М | Р | Р | Р | Р | — | — | — | Н | — | — | Н | — | Н | Н | Н | Н | Н | Н | Н | Н | Н |
| HS — | Р | Р | Р | Р | Р | Р | Р | Р | Р | ? | ? | ? | ? | ? | Н | ? | ? | ? | ? | ? | ? | ? |
| SO3 2- | Р | Р | Р | Р | Р | Н | Н | М | Н | ? | — | Н | ? | Н | Н | ? | М | М | — | Н | ? | ? |
| HSO3 — | Р | ? | Р | Р | Р | Р | Р | Р | Р | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? |
| SO4 2- | Р | Р | Р | Р | Р | Н | М | Р | Н | Р | Р | Р | Р | Р | Р | Р | Р | М | — | Н | Р | Р |
| HSO4 — | Р | Р | Р | Р | Р | Р | Р | Р | — | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | Н | ? | ? |
| NO3 — | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | — | Р |
| NO2 — | Р | Р | Р | Р | Р | Р | Р | Р | Р | ? | ? | ? | ? | Р | М | ? | ? | М | ? | ? | ? | ? |
| PO4 3- | Р | Н | Р | Р | — | Н | Н | Н | Н | Н | Н | Н | Н | Н | Н | Н | Н | Н | Н | Н | Н | Н |
| CO3 2- | Р | Р | Р | Р | Р | Н | Н | Н | Н | ? | ? | Н | ? | Н | Н | Н | Н | Н | ? | Н | ? | Н |
| CH3COO — | Р | Р | Р | Р | Р | Р | Р | Р | Р | — | Р | Р | — | Р | Р | Р | Р | Р | Р | Р | — | Р |
| SiO3 2- | Н | Н | Р | Р | ? | Н | Н | Н | Н | ? | ? | Н | ? | ? | ? | Н | Н | ? | ? | Н | ? | ? |
| Растворимые (>1%) | Нерастворимые ( Спасибо! Ваша заявка отправлена, преподаватель свяжется с вами в ближайшее время. Вы можете также связаться с преподавателем напрямую: 8(906)72 3-11-5 2
Скопируйте эту ссылку, чтобы разместить результат запроса » » на другом сайте. Изображение вещества/реакции можно сохранить или скопировать, кликнув по нему правой кнопкой мыши. Если вы считаете, что результат запроса » » содержит ошибку, нажмите на кнопку «Отправить». Этим вы поможете сделать сайт лучше. К сожалению, регистрация на сайте пока недоступна. На сайте есть сноски двух типов: Подсказки — помогают вспомнить определения терминов или поясняют информацию, которая может быть сложна для начинающего. Дополнительная информация — такие сноски содержат примечания или уточнения, выходящие за рамки базовой школьной химии, нужны для углубленного изучения. Здесь вы можете выбрать параметры отображения органических соединений. Аминокислоты. Общие пути обмена аминокислот. Синтез мочевины.» data-shape=»round» data-use-links data-color-scheme=»normal» data-direction=»horizontal» data-services=»messenger,vkontakte,facebook,odnoklassniki,telegram,twitter,viber,whatsapp,moimir,lj,blogger»> Аминокислоты. Общие пути обмена аминокислот. Биосинтез мочевины. Аминокислоты – органические соединения, содержащие –СООН и -NH2 в α-положении. Почти все аминокислоты имеют хиральный атом и обладают оптической изомерией. У человека присутствуют L-аминокислоты Аминокислоты. Физико-химические свойства К наиболее важным общим реакциям аминокислот относятся реакции:
Специфические реакции аминокислот связаны с наличием функциональных групп в радикале (окислительно-восстановительные реакции цис). Аминокислоты. Классификация. Классификация аминокислот по природе радикала:
Классификация аминокислот по количеству карбоксильных и аминогрупп:
Классификация аминокислот по функциональным группам в радикале:
Классификация аминокислот по способности к синтезу:
Аминокислоты. Функции
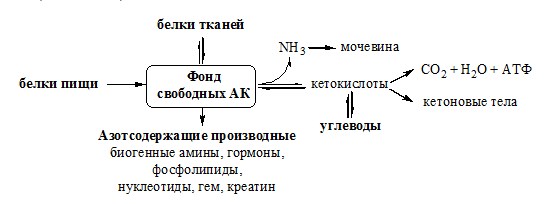
ПУТИ ОБРАЗОВАНИЯ ПУЛА АМИНОКИСЛОТ В КРОВИ Большая часть аминокислот организма человека, примерно 15кг, входит в состав белков. Фонд свободных аминокислот организма составляет примерно 35г.
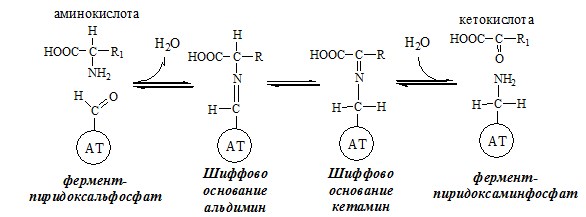
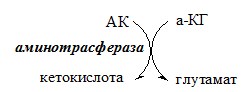
ОБЩИЕ РЕАКЦИИ ОБМЕНА АМИНОКИСЛОТ Аминокислоты, появившиеся в организме, включаются в общие и специфические реакции обмена. ТРАНСАМИНИРОВАНИЕ (ПЕРЕАМИНИРОВАНИЕ) АМИНОКИСЛОТ Трансаминирование — реакция переноса α-аминогруппы с аминокислоты на α-кетокислоту, в результате чего образуются новая α-кетокислота и новая аминокислота. Процесс трансаминирования легко обратим, при нем общее количество аминокислот в клетке не меняется.
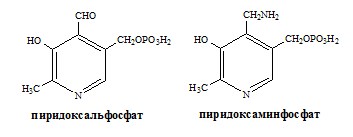
Реакции катализируют аминотрансферазы, коферментом которых служит пиридоксальфосфат (ПФ) — производное витамина В6 (пиридоксина).
Механизм переаминирования
Вначале, аминоксилота передает свою аминогруппу на пиродоксальфосфат. Аминокислота при этом превращается в кетокислоту, а пиродоксальфосфат – в пиридоксаминфосфат.
Органоспецифичные аминотрансферазы АЛТ и АСТ Чаще всего в трансаминировании участвуют аминокислоты и кетокислоты, которых много в организме — глу, ала, асп, α-КГ, ПВК и ЩУК. Основным донором аминогруппы служит глу, а кетогруппы – α-КГ. Биологическое значение трансаминирования ДЕЗАМИНИРОВАНИЕ АМИНОКИСЛОТ Дезаминирование аминокислот — реакция отщепления α-аминогруппы от аминокислоты, в результате чего образуется соответствующая α-кетокислота и выделяется молекула аммиака. Прямое дезаминирование аминокислоты Существует 5 видов прямого дезаминирования аминокислот:
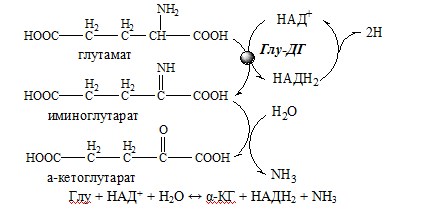
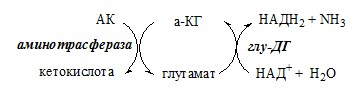
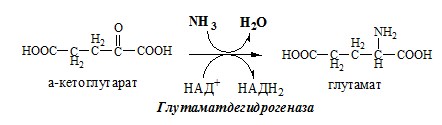
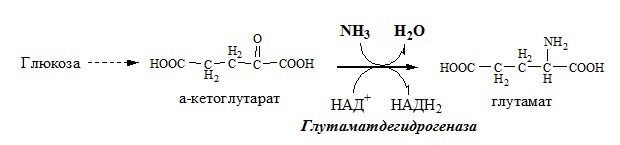
Окислительное дезаминирование – самый активный вид прямого дезаминирования аминокислот. 1. Глутаматдегидрогеназа (глу-ДГ) – олигомер, состоящий из 6 субъединиц (молекулярная масса 312 кД), содержит кофермент НАД+. Глу-ДГ катализирует обратимое дезаминирование глу, очень активна в митохондриях клеток практически всех органов, кроме мышц. Глу-ДГ аллостерически ингибируют АТФ, ГТФ, НАДH2, активирует избыток АДФ. Индуцируется Глу-ДГ стероидными гормонами (кортизолом).
Глу + НАД+ + Н2О ↔ α-КГ + НАДН2 + NH3 2. Оксидаза L-аминокислот
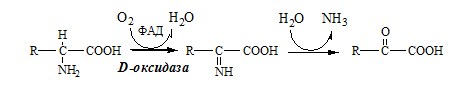
Оксидаза L-АК имеет кофермент ФМН. Т.к. оптимум рН оксидазы L-АК равен 10,0, активность фермента очень низка и вклад ее в дезаминирование незначителен. 3. Оксидаза D-аминокислот
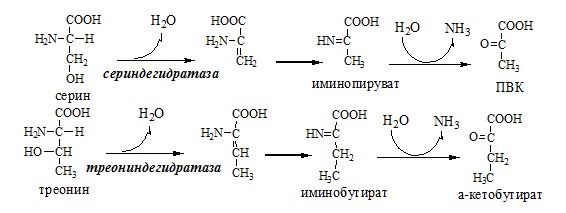
Неокислительное дезаминирование В печени человека присутствуют специфические пиридоксальфосфатзависимые ферменты сериндегидратаза, треониндегидратаза, катализирующие реакции неокислительного дезаминирования аминокислот серина и треонина.
Внутримолекулярное дезаминирование
Непрямое дезаминирование (трансдезаминирование) аминокислот Непрямое дезаминирование в печени
Обе стадии непрямого дезаминирования обратимы, что обеспечивает как катаболизм аминокислот, так и возможность образования практически любой АК из соответствующей α-кетокислоты. Непрямое дезаминирование в мышцах (и нервной ткани)
Можно выделить 4 стадии этого процесса:
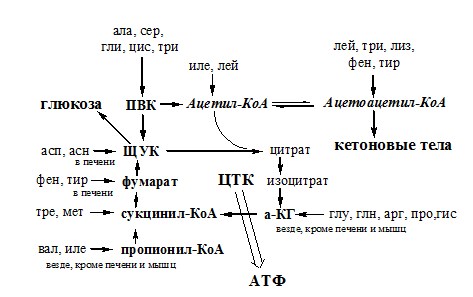
Этот путь дезаминирования преобладает в мышцах при интенсивной работе, в результате которой накапливается молочная кислота. Выделяющийся аммиак предотвращает закисление среды в клетках, вызванное образованием лактата. Пути обмена безазотистого остатка аминокислот За сутки у человека распадаются примерно 100г АК. Катаболизм всех АК сводится к образованию шести веществ, вступающих в общий путь катаболизма: ПВК, ацетил-КоА, α-кетоглутарат, сукцинил-КоА, фумарат и ЩУК. Эти вещества окисляются в ЦТК для образования АТФ или используются для синтеза глюкозы и кетоновых тел.
Гликогенные аминокислоты – АК, которые превращаются в ПВК и промежуточные продукты ЦТК (а-КГ, сукцинил-КоА, фумарат, ЩУК). Они через ЩУК, используются в глюконеогенезе (ала, асн, асп, гли, глу, глн, про, сер, цис, арг, гис, вал, мет, тре). ОБМЕН АММИАКА Аммиак в организме образуется:
Концентрация аммиака Токсичность аммиака Механизм токсического действия аммиака:
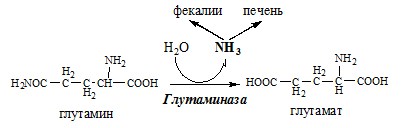
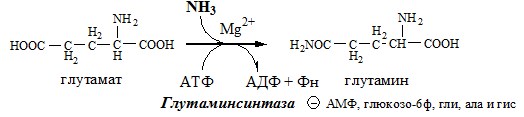
Связывание (обезвреживание) аммиака В связи с токсичностью аммиака в тканях происходит его связывание с образованием нетоксичных соединений – АК и мочевины. Процесс образования и обезвреживания аммиака регулируют в основном ферменты глутаматдегидрогеназа и глутаминсинтетаза. Обмен глутамата
Однако этот путь обезвреживания аммиака в тканях используется слабо, так как глутаматдегидрогеназа катализирует преимущественно реакцию дезаминирования глутамата. Хотя, если учитывать последующее образование глутамина, реакция выгодна для клеток, так как способствует связыванию сразу 2 молекул NH3. Обмен глутамина
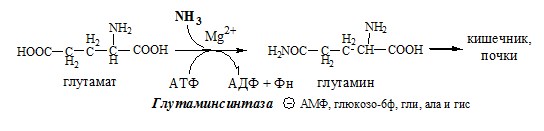
Глутаминсинтетаза находиться в митохондриях клеток, содержит кофактор — ионы Mg2+, является одним из основных регуляторных ферментов обмена АК. Она аллостерически ингибируется АМФ, глюкозо-6ф, гли, ала и гис.
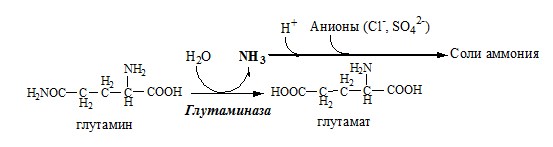
Аммиак с протонами и анионами образует соли аммония (0,5 г/сут), которые выделяются с мочой. Этот процесс используется для регуляции КОС и сохранения в организме важнейших катионов Na+ и К+. Глутаминаза почек значительно индуцируется при ацидозе, ингибируется при алкалозе.
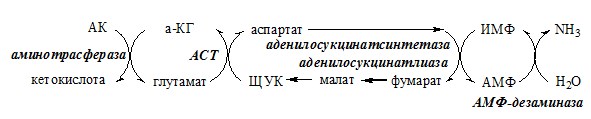
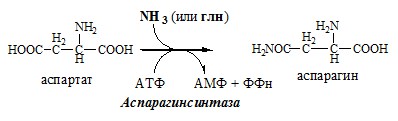
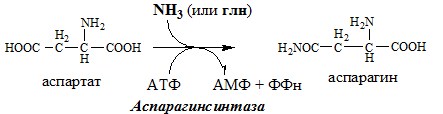
Образовавшийся аммиак поступает через воротную вену в печень или удаляется из организма с фекалиями. Обмен аспарагина Обезвреживание аммиака в тканях происходит незначительно при синтезе аспарагина под действием глутаминзависимой и аммиакзависимой аспарагинсинтетазы.
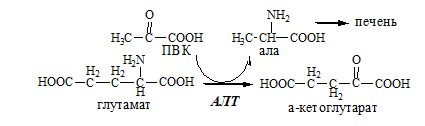
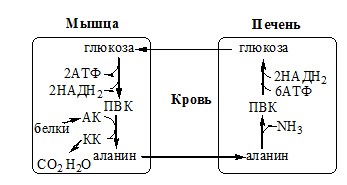
Первая функционирует в животных клетках, вторая преобладает в бактериальных клетках, но присутствует и у животных. Обмен аланина Из мышц и кишечника избыток аминого азота выводится преимущественно в виде аланина.
Глутамат подвергается трансаминированию с ПВК с образованием аланина и α-кетоглутарата. Аланин поступает из кишечника в кровь воротной вены и поглощается печенью. Этот процесс называется глюкозо-аланиновый цикл:
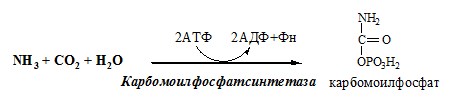
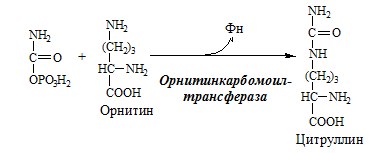
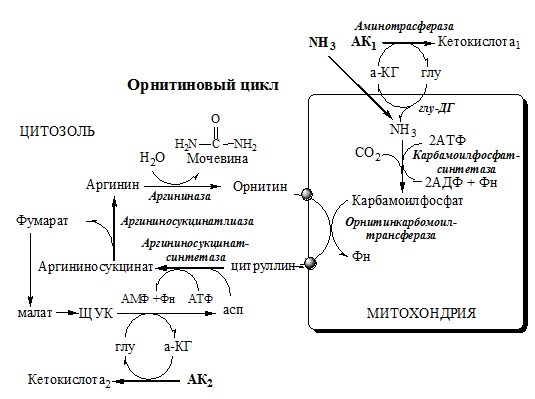
Он необходим, так как активность глу-ДГ в мышцах невелика и непрямое дезаминирование АК малоэффективно. ОРНИТИНОВЫЙ ЦИКЛ Большая часть свободного аммиака, а также аминного азота в составе (в основном глутамин, аланин) поступают в печень, где из них синтезируется нетоксичное и хорошо растворимое в воде соединение — мочевина. Мочевина является основной формой выведения азота из организма человека. Реакции орнитинового цикла
(Карбамоилфосфатсинтетаза II локализована в цитозоле клеток всех тканей и участвует в синтезе пиримидиновых нуклеотидов).
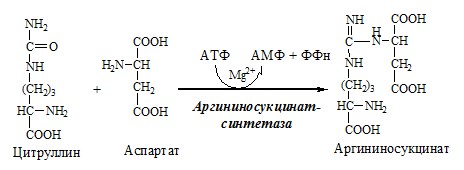
2. В цитозоле аргининосукцинатсинтетаза с затратой 1 АТФ (двух макроэргических связей) связывает цитруллин с аспартатом и образует аргининосукцинат (аргининоянтарная кислота). Фермент нуждается в Mg2+. Аспартат — источник второго атома азота мочевины.
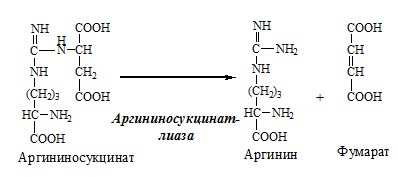
3. В цитозоле аргининосукцинатлиаза (аргининсукциназа) расщепляет аргининосукцинат на аргинин и фумарат (аминогруппа аспартата оказывается в аргинине).
4. В цитозоле аргиназа гидролизует аргинин на орнитин и мочевину. У аргиназы кофакторы ионы Са2+ или Мn2+, ингибиторы – высокие концентрации орнитина и лизина.
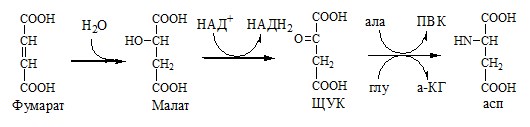
Образующийся орнитин взаимодействует с новой молекулой карбамоилфосфата, и цикл замыкается. Регенерация аспартата из фумарата Фумарат, образующийся в орнитиновом цикле, в цитозоле превращается в ЩУК, который переаминируется с аланином или глутаматом с образованием аспартата. Аланин поступает главным образом из мышц и клеток кишечника:
Малат может направиться в митохондрии и включиться в ЦТК.
Энергетический баланс орнитинового цикла Энергозатраты при этом частично компенсируются:
Орнитиновый цикл в печени выполняет 2 функции:
Полный набор ферментов орнитинового цикла есть только в гепатоцитах. Отдельные же ферменты орнитинового цикла обнаруживаются в разных тканях. В энтероцитах, есть карбамоилфосфатсинтетаза I и орнитинкарбамоилтрансфераза, следовательно, может синтезироваться цитруллин. В почках есть аргининосукцинатсинтетаза и аргининосукцинатлиаза. Цитруллин, образовавшийся в энтероцитах, может поступать в почки и превращаться там в аргинин, который переносится в печень и гидролизуется аргиназой. Активность этих рассеянных по разным органам ферментов значительно ниже, чем в печени. Выделение азота из организма ГИПЕРАММОНИЕМИЯ Нарушение реакций обезвреживания аммиака может вызвать повышение содержания аммиака в крови — гипераммониемию, что оказывает токсическое действие на организм. Причинами гипераммониемии могут быть: Известны пять наследственных заболеваний, обусловленные дефектом пяти ферментов орнитинового цикла. Наследственные нарушения орнитинового цикла и их основные проявления Гиперам-мониемия, тип I В течение 24-48 ч после рождения кома, смерть Гиперам-мониемия, тип II Сцепленный с Х-хромосомой Гипотония, снижение толерантности к белкам Гипераммониемия тяжёлая у новорождённых. У взрослых — после белковой нагрузки Гипераммониемия, атаксия, судороги, выпадение волос Аргини-носукцинат NH 3 Аргини-носукци-нат, Глн, Ала, Лиз Apг Снижение активности какого-либо фермента синтеза мочевины приводит к накоплению в крови субстрата данного фермента и его предшественников. Гипераммониемия сопровождается появлением следующих симптомов:
В тяжёлых случаях развивается кома с летальным исходом. Лечение больных с различными дефектами орнитинового цикла в основном направлено на снижение концентрации аммиака в крови за счёт малобелковой диеты, введения кетоаналогов АК в рацион и стимуляцию выведения аммиака в обход нарушенных реакций:
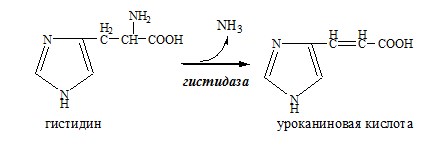
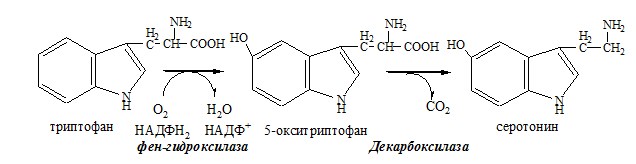
Обмен аминокислот и аммиака между тканями Печень Мышцы Кишечник Мозг Почки ДЕКАРБОКСИЛИРОВАНИЕ АМИНОКИСЛОТ И ИХ ПРОИЗВОДНЫХ Некоторые аминокислоты и их производные могут подвергаться декарбоксилированию – отщеплению α-карбоксильной группы. У млекопитающих декарбоксилируются: три, тир, вал, гис, глу, цис, арг, орнитин, SAM, ДОФА, 5-окситриптофан и т.д. Реакцию необратимо катализируют декарбоксилазы, которые содержат в активном центре пиридоксальфосфат. Механизм реакции похож на реакцию переаминирования. Серотонин
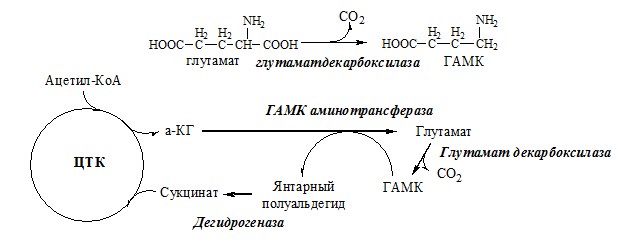
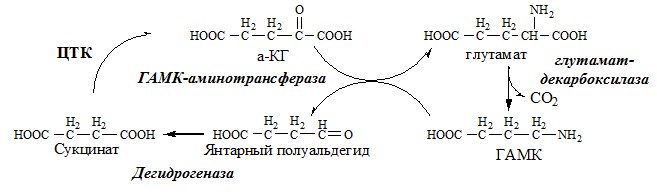
ГАМК
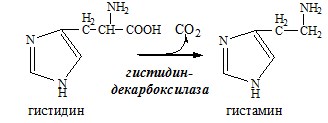
ГАМК – тормозной нейромедиатор (повышает проницаемость постсинаптических мембран для К+), повышает дыхательную активность нервной ткани, улучшает кровоснабжение головного мозга. Гистамин
Гистамин – медиатор воспаления, аллергических реакций, пищеварительный гормон:
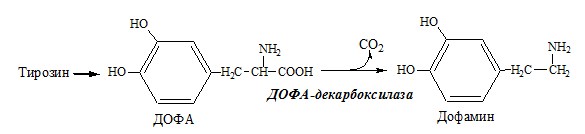
Дофамин
Дофамин – нейромедиатор среднего отдела мозга. Аминокислоты. Специфические пути обмена аминокислот ФОЛИЕВАЯ КИСЛОТА Значительную роль в обмене ряда АК, синтезе некоторых сложных липидов, нейромедиаторов, гормонов и ряда других веществ играют производные фолиевой кислоты.
Активная форма фолиевой кислоты – ТГФК. Она образуется в печени при восстановлении фолиевой кислоты с участием фолатредуктазы и дигидрофолатредуктазы, коферментом которых служит НАДФН2.
Образование одноуглеродных фрагментов, их взаимопревращения ТГФК принимает от АК одноуглеродные фрагменты: серин и глицин дают метиленовый фрагмент (-СН2-), гистидин – формимино- и формильный фрагменты.
Затем ТГФК отдает одноуглеродные фрагменты на:
Недостаточность фолиевой кислоты Гиповитаминоз фолиевой кислоты приводит к:
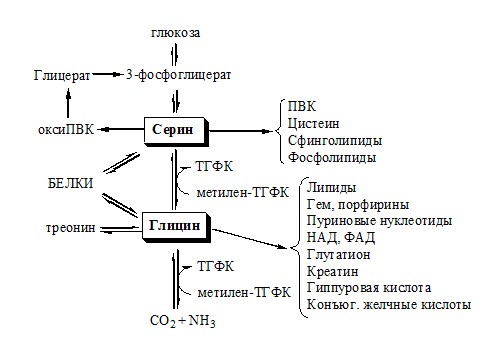
КОБАЛАМИН (В12) В12 синтезируется только микроорганизмами, им богаты печень, почки. Активные формы кобаламина – метилкобаламин (цитоплазма) и дезоксиаденозилкобаламин (митохондрии). Кобаламин участвует: Недостаточность В12 Гиповитаминоз В12 сопровождается: ОБМЕН СЕРИНА И ГЛИЦИНА Серин и глицин – заменимые аминокислоты.
Схема путей обмена серина и глицина
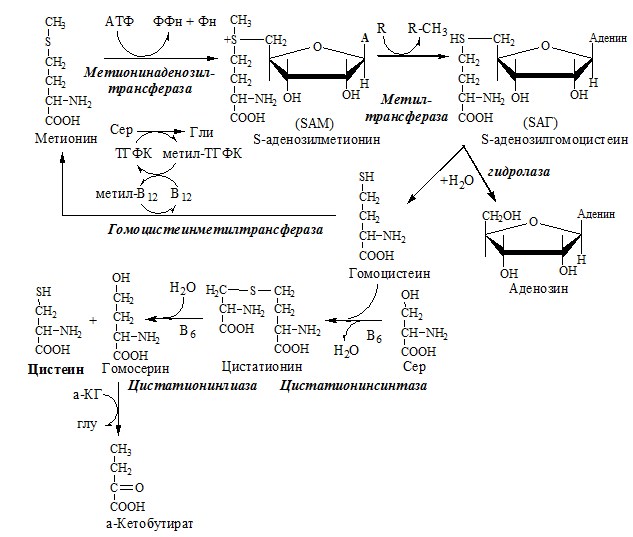
Глицин — важнейший (после ГАМК) тормозной нейромедиатор в спинном мозге, промежуточном мозге и некоторых отделах головного мозга. Наследственные нарушения обмена глицина Известно несколько заболеваний, связанных с нарушениями обмена глицина. В их основе лежит недостаточность ферментов или дефект системы транспорта этой АК. В состав белков человека входят 2 АК, содержащие серу, — метионин и цистеин. Эти аминокислоты метаболически тесно связаны между собой. МЕТИОНИН Метионин — незаменимая аминокислота, может регенерировать из гомоцистеина с участием серина и глицина. Метионин: Образование S-аденозилметионина Ресинтез метионина, роль ТГФК и витамина В12.
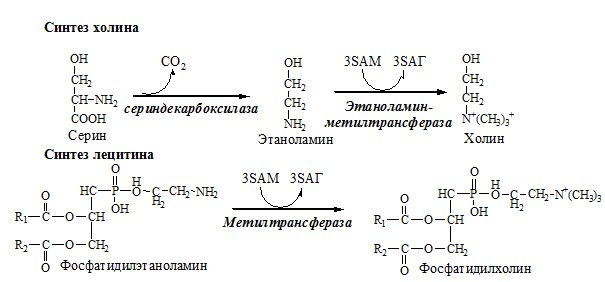
Реакции трансметилирования с участием S-аденозилметионина Синтез холина и синтез лецитина
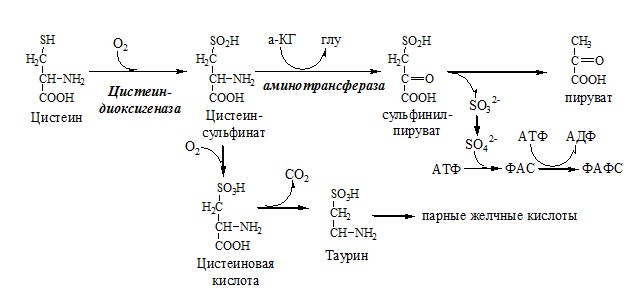
Аналогично синтезируются: ЦИСТЕИН Цистеин – серосодержащая условнозаменимая АК. Синтезируется из незаменимого метионина и заменимого серина. Цистеин:
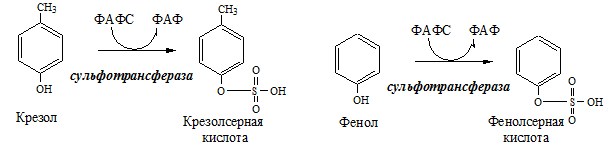
Образование сульфат-иона, его утилизация (образование ФАФС).
ФАФС используется:
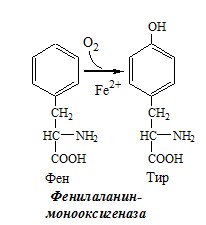
2. В синтезе гликозаминогликанов (сульфирование ОН групп производных глюкозы, галактозы сульфотрансферазой). ФЕНИЛАЛАНИН Фенилаланин — незаменимая АК, которая содержится в достаточных количествах в пищевых продуктах. Фенилаланин идет в основном на синтез белков и тирозина.
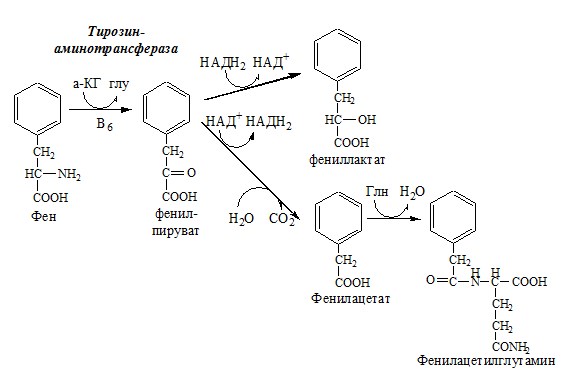
Фенилкетонурия Классическая фенилкетонурия— наследственное заболевание, связанное с мутациями в гене фенилаланингидроксилазы (частота 1:10000 новорождённых), которые приводят к снижению активности фермента или полной его инактивации. Проявления фенилкетонурии: Проявления фенилкетонурии связаны с токсическим действием на клетки мозга высоких концентраций фенилаланина, фенилпирувата, фениллактата. Большие концентрации фенилаланина ограничивают транспорт тирозина и триптофана через гематоэнцефалический барьер и тормозят синтез нейромедиаторов (дофамина, норадреналина, серотонина).
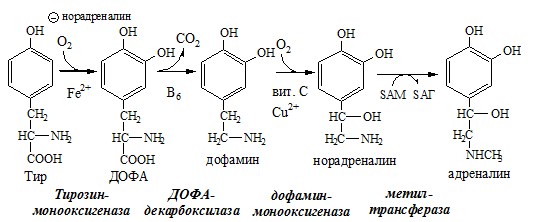
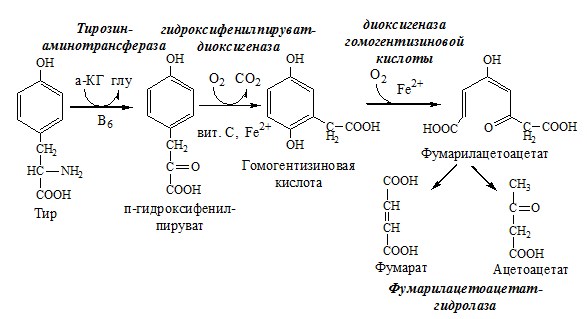
ТИРОЗИН Тирозин — условно заменимая АК, образуется из незаменимого фенилаланина. Содержание тир в пищевых белках достаточно велико. Обмен тирозина зависит от типа тканей. 1. Обмен тирозина в надпочечниках и нервной ткани
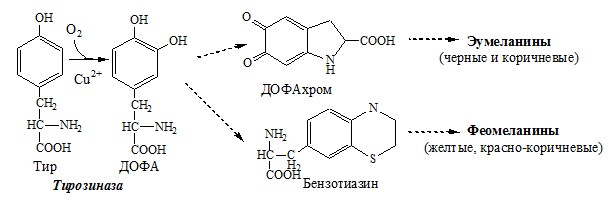
Тирозингидроксилаза (тирозинмонооксигеназа) Fe2+ -зависимый фермент, в качестве кофермента использующий Н4БП. Ее ингибирует норадреналин. Болезнь Паркинсона 2. Обмен тирозина в меланоцитах В пигментных клетках (меланоцитах) обмен тирозин идет по меланиновому пути. Из тирозина синтезируются пигменты — меланины 2 типов: эумеланины и феомеланины. Эумеланины (чёрного и коричневого цвета) — нерастворимые высокомолекулярные полимеры 5,6-дигидроксииндола. Феомеланины — жёлтые или красновато-коричневые полимеры, растворимые в разбавленных щелочах.
Альбинизм 3. Превращение тирозина в щитовидной железе 4. Катаболизм тирозина в печени
Фумарат может окисляться до СО2 и Н2О или использоваться для глюконеогенеза. Ацетоацетат — кетоновое тело, окисляемое до СО2 и Н2О с выделением энергии. Алкаптонурия («чёрная моча») Тирозинемии Различают 3 типа тирозинемии.
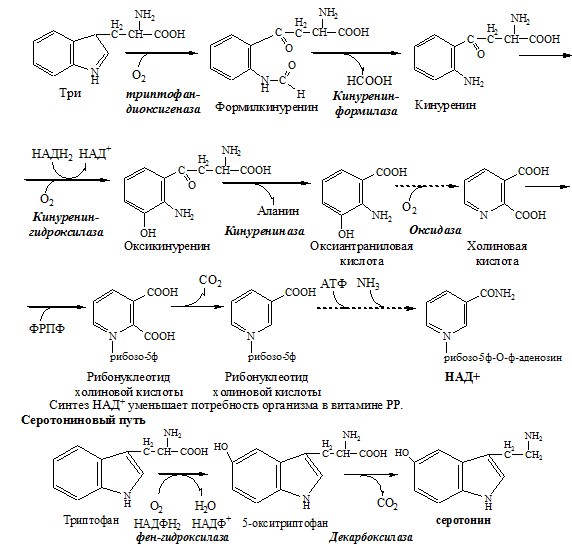
ТРИПТОФАН Триптофан – незаменимая АК. В физиологических условиях >95% триптофана метаболизирует по кинурениновому пути и 1% по серотониновому пути. Схема кинуренинового пути
Серотонин образуется в надпочечниках, ЦНС и тучных клетках. Образование и использование в организме ГАМК и ГОМК. Антиоксидантные, антигипоксические и адаптогенные свойства Глу, Асп, их клиническое применение. ГЛУТАМАТ Синтез глутамата
2). В реакция переаминирования с участием аминотрансфераз:
Использование глутамата
Глутамат содержится в больших количествах в головном мозге, где выполняет разнообразные функции:
Нарушение обмена глутамата приводит к целому ряду патологических нарушений ЦНС: эпилепсии, расстройствах вестибулярной системы, ишемии и др. Глутамат и его аналоги используют как лекарственные средства при хронической недостаточности аминокислотного обмена, вегетососудистой дистонии, эпилепсии (в качестве предшественника ГАМК — тормозного медиатора). ГЛУТАМИН Синтез глутамина
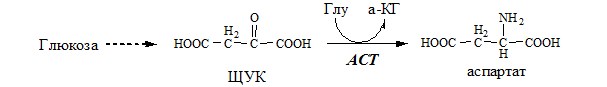
Использование глутамина АСПАРТАТ Синтез аспартата
Использование аспартата АСПАРАГИН Синтез аспарагина
Использование аспарагина источники: http://acetyl.ru/o/a21k2w.php http://farmf.ru/lekcii/aminokisloty-obshhie-puti-obmena-aminokislot-sintez-mocheviny/ |