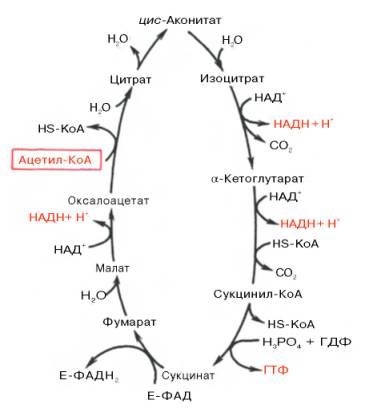
Цикл трикарбоновых кислот Кребса реакции (Схема)
Цикл трикарбоновых кислот (Цикл Кребса или цикл лимонной кислоты) — центральный процесс метаболизма. Суть процесса состоит в том, что первоначально пировиноградная кислота соединяется со сложным органическим веществом — коэнзимом А. При этом сразу отщепляется СО2, образуется НАД • Н и комплекс СН3СО—5—коэнзим А (ацетил-коэнзим А). Этот комплекс реагирует со щавелево-уксусной кислотой с образованием кислоты лимонной. Коэнзим А освобождается, а лимонная кислота подвергается последовательным превращениям в цикле Кребса (8 реакций). При этом получается 2 молекулы СО2, восстанавливается молекула щавелево-уксусной кислоты, а выделяющаяся энергия запасается в виде АТФ. В пересчете на одну молекулу глюкозы энергетический выход цикла Кребса составляет 30 молекул АТФ. Таким образом, суммарный энергетический выход всего процесса дыхания составляет 38 молекул АТФ на одну молекулу глюкозы. Часть образующейся энергии выделяется в виде тепла (44,6%). Цикл Кребса идет только в присутствии необходимого количества кислорода. Этот цикл открыл Г. Кребс, за которое он в 1953 году получил Нобелевскую премию.
Схема цикл трикарбоновых кислот, цикл Кребса

ATP (АТФ) — это аденозинтрифосфорная кислота, универсальный источник и переносчик энергии
NAD (НАД) — никотинамидадениндинуклеотидфосфата, кофермент
NADH (НАД•Н) — востановленная форма NAD
Ацетил-КоА — сложное органическое вещество ацетил-коэнзим А (СН3СО—S)
Пируват — это соли пировиноградной кислоты
Таблица реакции цикла трикарбоновых кислот Кребса
Цикл Кребса это аэробный процесс (проходит с участием кислорода) и состоит из восьми стадий (процессов).
Реакции цикла трикарбоновых кислот (цикла Кребса)
Описание и ход реакция цикла
Конденсация ацетил-КоА с оксалоацетатом
Конденсация ацетил-СоА с оксалоацетатом, в результате чего образуется цитрат (лимонная кислота), а кофермент А освобождается. Реакция катализируется цитратсинтазой, которая является одним из регуляторных ферментов, лимитирующих скорость цикла Кребса.

Изомеризация цитрата в изоцитрат
Превращение цитрата в изоцитрат (изолимонная кислота) с участием аконитат-гидратазы (сложного фермента, содержащего Fe 2+ и кислотолабильные атомы серы, образующие железо-серные центры) через промежуточную стадию цис-аконитата, связанного с ферментом.

Окислительное декарбоксилирование изоцитрата
Дегидрирование (так называется удаление из молекулы атомов водорода) цитрата с образованием a-кетоглутарата и СO2 при участии изоцитратдегидрогеназы, которая функционирует при наличии Mg 2+ и Мn 2+ .

Окислительное декарбоксилирование а-кетоглутарата
Реакция окислительного декарбоксилирования a-кетоглутарата до высокоэнергетического сукцинил-СоА. Реакция катализируется a-кетоглутаратдегидрогеназным комплексом (Mg 2+ ), который похож на пируватдегидрогеназный комплекс.

коферменты: ТПФ, липоевая кислота, ФАД, HS-KoA, НАД+
На этом этапе цикла Кребса происходит превращение сукцинил-СоА под влиянием сукцинил-СоА-синтетазы в сукцинат (янтарная кислота) с отщеплением СоА. Эта реакция сопряжена с образованием гуанозинтрифосфата (GTP) из GDP и фосфата и также катализируется указанным ферментом.

Катализируемое сукцинатдегидрогеназой, содержащей ковалентно связанный FAD и два железо-серных центра, дегидрирование сукцината с образованием фумарата (фумаровая кислота).

Эта реакция цикла трикарбоновых кислот представляет обратимую гидратацию фумарата с образованием L-малата (яблочная кислота), катализируемая фумарат-гидратазой или фумаразой.

Катализируемое NAD-зависимой L-малатдегидрогеназой дегидрирование L-малата с образованием оксалоацетата. Эта реакция замыкает цикл Кребса и поставляет оксалоацетат (щавелевоянтарная кислота) для нового цикла. Большинство реакций цикла Кребса обратимы.

_______________
Источник информации:
1. Биология для поступающих в вузы / Г.Л. Билич, В.А. Крыжановский. — 2008.
2. Биохимия в схемах и таблицах / И. В. Семак — Минск — 2011.
Реакции цикла Кребса

Продолжаем разбирать цикл Кребса. В прошлой статье я рассказывал о том, что это вообще такое, для чего цикл Кребса нужен и какое место в метаболизме он занимает. Теперь давайте приступим к самим реакциям этого цикла.
Сразу оговорюсь — лично для меня заучивание реакций было совершенно бессмысленным занятием до того, пока я не разобрал вышеуказанные вопросы. Но если вы уже разобрались с теорией, предлагаю перейти к практике.
Вы можете увидеть множество способов написания цикла Кребса. Чаще всего встречаются варианты вроде этого:
Но мне удобнее всего показался способ написания реакций из старого доброго учебника по биохимии от авторов Берёзова Т.Т. и Коровкина Б.В.
Первая реакция
Уже знакомые нам Ацетил-КоА и Оксалоацетат соединяются и превращаются в цитрат, то есть в лимонную кислоту.
Вторая реакция
Теперь берём лимонную кислоту и превращаем её изолимонную кислоту. Другое название этого вещества — изоцитрат.
На самом деле, эта реакция идёт несколько сложнее, через промежуточную стадию — образование цис-аконитовой кислоты. Но я решил упростить, чтобы вы получше запомнили. При необходимости вы сможете добавить сюда недостающую ступень, если будете помнить всё остальное.
По сути, две функциональные группы просто поменялись местами.
Третья реакция
Итак, у нас получилась изолимонная кислота. Теперь её нужно декарбоксилировать (то есть отщипнуть COOH) и дегидрировать (то есть отщипнуть H) . Получившееся вещество — это a-кетоглутарат.
Эта реакция примечательна тем, что здесь образуется комплекс HAДH2. Это значит, что переносчик НАД подхватывает водород, чтобы запустить дыхательную цепь.
Мне нравится вариант реакций Цикла Кребса в учебнике Берёзова и Коровкина именно тем, что сразу отлично видно атомы и функциональные группы, которые участвуют в реакциях.
Четвёртая реакция
Едем далее. Берём a-кетоглутарат из прошлой реакции и декарбоксилируем на сей раз его. Как видите, в этой же реакции к a-кетоглутарату присоединяется коэнзим-А.
Снова как часы работает никотинАмидАденинДинуклеотид, то есть НАД. Это славный переносчик появляется здесь, как и в прошлом шаге, чтобы захватить водород и унести его в дыхательную цепь.
Кстати, получившееся вещество — сукцинил-КоА, не должно вас пугать. Сукцинат — это другое название янтарной кислоты, хорошо знакомой вам со времён биоорганической химии. Сукцинил-Коа — это соединение янтарной кислоты с коэнзимом-А. Можно сказать, что это эфир янтарной кислоты.
Пятая реакция
В прошлом шаге мы говорили, что сукцинил-КоА — это эфир янтарной кислоты. А теперь мы получим саму янтарную кислоту, то есть сукцинат, из сукцинила-КоА. Крайне важный момент: именно в этой реакции происходит субстратное фосфорилирование.
Фосфорилирование вообще (оно бывает окислительное и субстратное) — это добавление фосфорной группы PO3 к ГДФ или АТФ, чтобы получить полноценный ГТФ, или соответственно, АТФ. Субстратное отличается тем, что эта самая фосфорная группа отрывается от какого-либо вещества, её содержащую. Ну проще говоря, она переносится с СУБСТРАТА на ГДФ или АДФ. Поэтому и называется — «субстратное фосфорилирование».
Ещё раз: на момент начала субстратного фосфорилирования у нас имеется дифосфатная молекула — гуанозинДифосфат или аденозинДифосфат. Фосфорилирование заключается в том, что молекула с двумя остатками фосфорной кислоты — ГДФ или АДФ «достраивается» до молекулы с тремя остатками фосфорной кислоты, чтобы получились гуанозинТРИфосфат или аденозинТРИфосфат. Этот процесс происходит во время превращения сукцинила-КоА в сукцинат (то есть, в янтарную кислоту).
На схеме вы можете увидеть буквы Ф (н). Это значит «неорганический фосфат». Неорганический фосфат переходит от субстрата на ГДФ, чтобы в продуктах реакции был хороший, полноценный ГТФ. Теперь давайте посмотрим на саму реакцию:
Шестая реакция
Следующее превращение. На сей раз янтарная кислота, которую мы получили в прошлом этапе, превратится в фумарат, обратите внимание на новую двойную связь.
На схеме отлично видно, как в реакции участвует ФАД: этот неутомимый переносчик протонов и электронов подхватывает водород и утаскивает его непосредственно в дыхательную цепь.
Седьмая реакция
Мы уже на финишной прямой. Предпоследняя стадия Цикла Кребса — это реакция превращения фумарата в L-малат. L-малат — это другое название L-яблочной кислоты, знакомой ещё с курса биоорганической химии.
Если вы посмотрите на саму реакцию, вы увидите, что, во-первых, она проходит в обе стороны, а во-вторых, её суть — гидратирование. То есть фумарат просто присоединяет к себе молекулу воды, в итоге получается L-яблочная кислота.
Восьмая реакция
Последняя реакция Цикла Кребса — это окисление L-яблочной кислоты до оксалоацетата, то есть до щавелевоуксусной кислоты. Как вы поняли, «оксалоацетат» и «щавелевоуксусная кислота» — это синонимы. Вы, наверное, помните, что щавелевоуксусная кислота является компонентом первой реакции цикла Кребса.
Здесь же отмечаем особенность реакции: образование НАДH2, который понесёт электроны в дыхательную цепь. Не забудьте также реакции 3,4 и 6, там также образуются переносчики электронов и протонов для дыхательной цепи.
Как видите, я специально выделил красным цветом реакции, в ходе которых образуются НАДH и ФАДH2. Это очень важные вещества для дыхательной цепи. Зелёным я выделил реакцию, в рамках которой происходит субстратное фосфорилирование, и получается ГТФ.
Как это всё запомнить?
На самом деле, не так уж и сложно. Полностью прочитав две моих статьи, а также ваш учебник и лекции, вам нужно просто потренироваться писать эти реакции. Я рекомендую запомнить цикл Кребса блоками по 4 реакции. Напишите эти 4 реакции несколько раз, для каждой подбирая ассоциацию, подходящую именно вашей памяти.
Например, мне сразу очень легко запомнилась вторая реакция, в которой из лимонной кислоты (она, думаю, всем знакома с детства) образуется изолимонная кислота.
Вы можете так же использовать мнемонические запоминалки, такие как: «Целый Ананас И Кусочек Суфле Сегодня Фактически Мой Обед, что соответствует ряду — цитрат, цис-аконитат, изоцитрат, альфа-кетоглутарат, сукцинил-CoA, сукцинат, фумарат, малат, оксалоацетат». Есть ещё куча подобных.
Но, если честно, мне не нравились такие стихи практически никогда. По-моему, проще запомнить саму последовательность реакций. Мне отлично помогло разделение цикла Кребса на две части, каждую из которых я тренировался писать по несколько раз в час. Как правило, это происходило на парах вроде психологии или биоэтики. Это весьма удобно — не отвлекаясь от лекции, вы можете потратить буквально минутку, написав реакции так, как вы их запомнили, а затем сверить с правильным вариантом.
Кстати, в некоторых вузах на зачётах и экзаменах по биохимии преподаватели не требуют знания самих реакций. Нужно знать только что такое цикл Кребса, где он происходит, в чём его особенности и значение, и, разумеется, саму цепочку превращений. Только цепочку можно называть без формул, используя лишь названия веществ. Такой подход не лишён смысла, на мой взгляд.
Надеюсь, моё руководство по циклу трикарбоновых кислот вам помогло. А я хочу напомнить, что эти две статьи не являются полноценной заменой вашим лекциям и учебникам. Я написал их лишь для того, чтобы вы примерно понимали, что такое цикл Кребса. Если вы вдруг увидели какую-то ошибку в моём руководстве, пожалуйста, отпишитесь о ней в комментариях. Спасибо за внимание!
ЦИКЛ ТРИКАРБОНОВЫХ КИСЛОТ (ЦИКЛ КРЕБСА)
Цикл трикарбоновых кислот впервые был открыт английским биохимиком Г. Кребсом. Он первым постулировал значение данного цикла для полного сгорания пирувата, главным источником которого является гликолитическое превращение углеводов. В дальнейшем было показано, что цикл трикарбо-новых кислот является тем центром, в котором сходятся практически все метаболические пути. Таким образом, цикл Кребса – общий конечный путь окисления ацетильных групп (в виде ацетил-КоА), в которые превращается в процессе катаболизма большая часть органических молекул, играющих роль «клеточного топлива»: углеводов, жирных кислот и аминокислот.
Образовавшийся в результате окислительного декарбоксилирования пирувата в митохондриях ацетил-КоА вступает в цикл Кребса. Данный цикл происходит в матриксе митохондрий и состоит из восьми последовательных реакций (рис. 10.9). Начинается цикл с присоединения ацетил-КоА к оксалоацетату и образования лимонной кислоты (цитрата). Затем лимонная кислота (шестиуглеродное соединение) путем ряда дегидрирований (отнятие водорода) и двух декарбоксилирований (отщепление СО2) теряет два углеродных атома и снова в цикле Кребса превращается в оксалоацетат (четырехуглеродное соединение), т.е. в результате полного оборота цикла одна молекула ацетил-КоА сгорает до СО2 и Н2О, а молекула окса-лоацетата регенерируется. Рассмотрим все восемь последовательных реакций (этапов) цикла Кребса.
Первая реакция катализируется ферментом цит-рат-синтазой, при этом ацетильная группа ацетил-КоА конденсируется с оксалоацетатом, в результате чего образуется лимонная кислота:
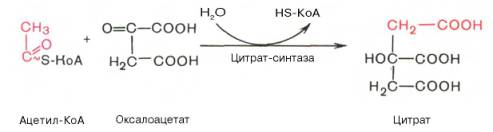
По-видимому, в данной реакции в качестве промежуточного продукта образуется связанный с ферментом цитрил-КоА. Затем последний самопроизвольно и необратимо гидролизуется с образованием цитрата и HS-KoA.
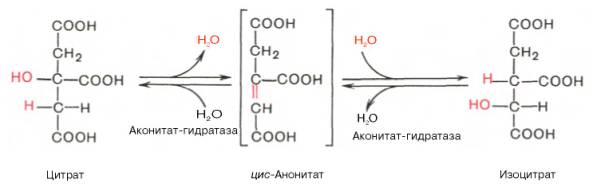
В результате второй реакции образовавшаяся лимонная кислота подвергается дегидратированию с образованием цис-аконитовой кислоты, которая, присоединяя молекулу воды, переходит в изолимонную кислоту (изоцитрат). Катализирует эти обратимые реакции гидратации–дегидратации фермент аконитатгидратаза (аконитаза). В результате происходит взаимоперемещение Н и ОН в молекуле цитрата:
Третья реакция, по-видимому, лимитирует скорость цикла Кребса. Изолимонная кислота дегидрируется в присутствии НАД-зависимой изо-цитратдегидрогеназы.
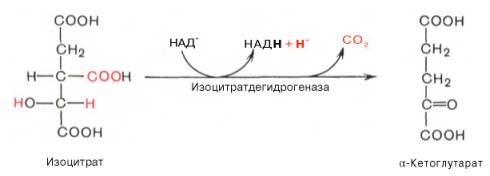
В ходе изоцитратдегидрогеназной реакции изолимонная кислота одновременно декарбоксилируется. НАД-зависимая изоцитратдегидрогеназа является аллостерическим ферментом, которому в качестве специфического активатора необходим АДФ. Кроме того, фермент для проявления своей активности нуждается в ионах Mg 2+ или Мn 2+ .
Во время четвертой реакции происходит окислительное декарбокси-лирование α-кетоглутаровой кислоты с образованием высокоэнергетического соединения сукцинил-КоА. Механизм этой реакции сходен с таковым реакции окислительного декарбоксилирования пирувата до ацетил-КоА, α-кетоглутаратдегидрогеназный комплекс напоминает по своей структуре пируватдегидрогеназный комплекс. Как в одном, так и в другом случае в реакции принимают участие 5 коферментов: ТПФ, амид липоевой кислоты, HS-KoA, ФАД и НАД + .
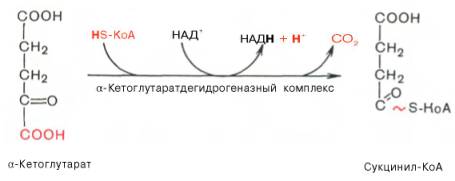
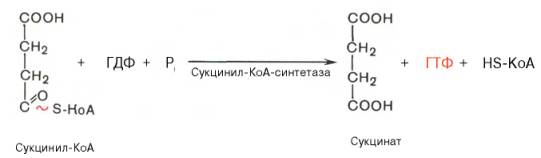
Пятая реакция катализируется ферментом сукцинил-КоА-синтета-зой. В ходе этой реакции сукцинил-КоА при участии ГТФ и неорганического фосфата превращается в янтарную кислоту (сукцинат). Одновременно происходит образование высокоэргической фосфатной связи ГТФ за счет высокоэргической тиоэфирной связи сукцинил-КоА:
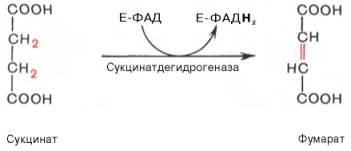
В результате шестой реакции сукцинат дегидрируется в фумаровую кислоту. Окисление сукцината катализируется сукцинатдегидрогеназой, в молекуле которой с белком прочно (ковалентно) связан кофермент ФАД. В свою очередь сукцинатдегидрогеназа прочно связана с внутренней ми-тохондриальной мембраной:
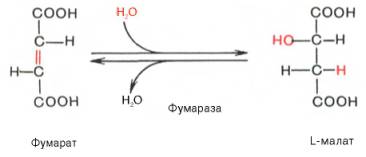
Седьмая реакция осуществляется под влиянием фермента фума-ратгидратазы (фумаразы). Образовавшаяся при этом фумаровая кислота гидратируется, продуктом реакции является яблочная кислота (малат). Следует отметить, что фумаратгидратаза обладает стереоспецифичностью (см. главу 4) – в ходе реакции образуется L-яблочная кислота:
Наконец, в ходе восьмой реакции цикла трикарбоновых кислот под влиянием митохондриальной НАД-зависимой малатдегидрогеназы происходит окисление L-малата в оксалоацетат:

Как видно, за один оборот цикла, состоящего из восьми ферментативных реакций, происходит полное окисление («сгорание») одной молекулы ацетил-КоА. Для непрерывной работы цикла необходимо постоянное поступление в систему ацетил-КоА, а коферменты (НАД + и ФАД), перешедшие в восстановленное состояние, должны снова и снова окисляться. Это окисление осуществляется в системе переносчиков электронов в дыхательной цепи (в цепи дыхательных ферментов), локализованной в мембране митохондрий. Образовавшийся ФАДН2 прочно связан с СДГ, поэтому он передает атомы водорода через KoQ. Освобождающаяся в результате окисления ацетил-КоА энергия в значительной мере сосредоточивается в макроэргических фосфатных связях АТФ. Из 4 пар атомов водорода 3 пары переносят НАДН на систему транспорта электронов; при этом в расчете на каждую пару в системе биологического окисления образуется 3 молекулы АТФ (в процессе сопряженного окислительного фосфорилирования), а всего, следовательно, 9 молекул АТФ (см. главу 9). Одна пара атомов от сукцинатдегидрогеназы-ФАДН2 попадает в систему транспорта электронов через KoQ, в результате образуется только 2 молекулы АТФ. В ходе цикла Кребса синтезируется также одна молекула ГТФ (субстратное фосфорилирование), что равносильно одной молекуле АТФ. Итак, при окислении одной молекулы ацетил-КоА в цикле Кребса и системе окислительного фосфорилирования может образоваться 12 молекул АТФ.
Если подсчитать полный энергетический эффект гликолитического расщепления глюкозы и последующего окисления двух образовавшихся молекул пирувата до СО2 и Н2О, то он окажется значительно большим.
Как отмечалось, одна молекула НАДН (3 молекулы АТФ) образуется при окислительном декарбоксилировании пирувата в ацетил-КоА. При расщеплении одной молекулы глюкозы образуется 2 молекулы пирувата, а при окислении их до 2 молекул ацетил-КоА и последующих 2 оборотов цикла трикарбоновых кислот синтезируется 30 молекул АТФ (следовательно, окисление молекулы пирувата до СО2 и Н2О дает 15 молекул АТФ). К этому количеству надо добавить 2 молекулы АТФ, образующиеся при аэробном гликолизе, и 6 молекул АТФ, синтезирующихся за счет окисления 2 молекул внемитохондриального НАДН, которые образуются при окислении 2 молекул глицеральдегид-3-фосфата в дегидрогеназной реакции гликолиза. Следовательно, при расщеплении в тканях одной молекулы глюкозы по уравнению С6Н12О6 + 6О2 —> 6СО2 + 6Н2О синтезируется 38 молекул АТФ. Несомненно, что в энергетическом отношении полное расщепление глюкозы является более эффективным процессом, чем анаэробный гликолиз.
Необходимо отметить, что образовавшиеся в процессе превращения глицеральдегид-3-фосфата 2 молекулы НАДН в дальнейшем при окислении могут давать не 6 молекул АТФ, а только 4. Дело в том, что сами молекулы внемитохондриального НАДН не способны проникать через мембрану внутрь митохондрий. Однако отдаваемые ими электроны могут включаться в митохондриальную цепь биологического окисления с помощью так называемого глицеролфосфатного челночного механизма (рис. 10.10). Ци-топлазматический НАДН сначала реагирует с цитоплазматическим ди-гидроксиацетонфосфатом, образуя глицерол-3-фосфат. Реакция катализи-
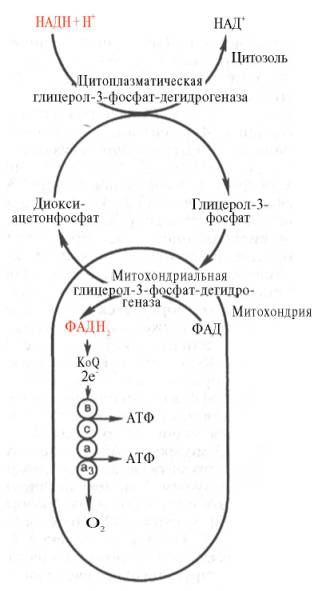
Рис. 10.10. Глицеролфосфатный челночный механизм. Объяснение в тексте.
руется НАД-зависимой цитоплазматической глицерол-3-фосфат-дегидроге-назой:
Дигидроксиацетонфосфат + НАДН + Н + Глицерол-3-фосфат + НАД + .
Образовавшийся глицерол-3-фосфат легко проникает через митохонд-риальную мембрану. Внутри митохондрии другая (митохондриальная) глицерол-3-фосфат-дегидрогеназа (флавиновый фермент) снова окисляет глицерол-3-фосфат до диоксиацетонфосфата:
Глицерол-3-фосфат + ФАД Диоксиацетонфосфат + ФАДН2.
Восстановленный флавопротеин (фермент-ФАДН2) вводит на уровне KoQ приобретенные им электроны в цепь биологического окисления и сопряженного с ним окислительного фосфорилирования, а диоксиаце-тонфосфат выходит из митохондрий в цитоплазму и может вновь взаимодействовать с цитоплазматическим НАДН + Н + . Таким образом, пара электронов (из одной молекулы цитоплазматического НАДН + Н + ), вводимая в дыхательную цепь с помощью глицеролфосфатного челночного механизма, дает не 3, а 2 АТФ.
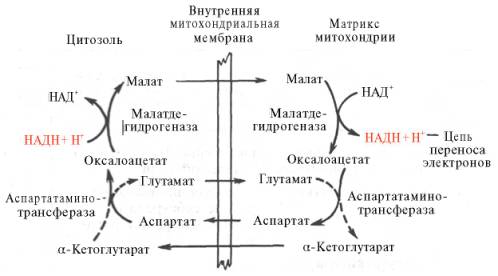
Рис. 10.11. Малат-аспартатная челночная система для переноса восстанавливающих эквивалентов от цитозольного НАДН в митохондриальный матрикс. Объяснение в тексте.
В дальнейшем было показано, что с помощью данного челночного механизма лишь в скелетных мышцах и мозге осуществляется перенос восстановленных эквивалентов от цитозольного НАДН + Н + в митохондрии.
В клетках печени, почек и сердца действует более сложная малат-ас-партатная челночная система. Действие такого челночного механизма становится возможным благодаря присутствию малатдегидрогеназы и ас-партатаминотрансферазы как в цитозоле, так и в митохондриях.
Установлено, что от цитозольного НАДН + Н + восстановленные эквиваленты сначала при участии фермента малатдегидрогеназы (рис. 10.11) переносятся на цитозольный оксалоацетат. В результате образуется малат, который с помощью системы, транспортирующей дикарбоновые кислоты, проходит через внутреннюю мембрану митохондрии в матрикс. Здесь малат окисляется в оксалоацетат, а матриксный НАД + восстанавливается в НАДН + Н + , который может теперь передавать свои электроны в цепь дыхательных ферментов, локализованную на внутренней мембране митохондрии. В свою очередь образовавшийся оксалоацетат в присутствии глутамата и фермента АсАТ вступает в реакцию трансаминирования. Образующиеся аспарат и α-кетоглутарат с помощью специальных транспортных систем способны проходить через мембрану митохондрий.
Транспортирование в цитозоле регенерирует оксалоацетат, что вызывает к действию следующий цикл. В целом процесс включает легкообратимые реакции, происходит без потребления энергии, «движущей силой» его является постоянное восстановление НАД + в цитозоле гли-церальдегид-3-фосфатом, образующимся при катаболизме глюкозы.
Итак, если функционирует малат-аспартатный механизм, то в результате полного окисления одной молекулы глюкозы может образоваться не 36, а 38 молекул АТФ (табл. 10.1).
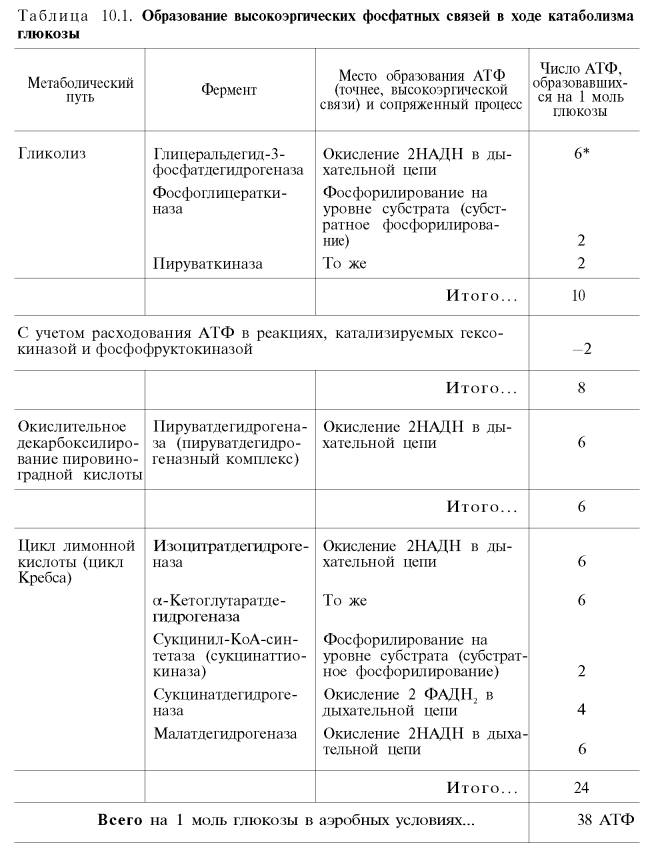
В табл. 10.1 приведены реакции, в которых происходит образование высокоэргических фосфатных связей в ходе катаболизма глюкозы, с указанием эффективности процесса в аэробных и анаэробных условиях.
http://medicine-boy.ru/reakcii_cikla_krebsa/
http://xumuk.ru/biologhim/151.html