ПЕНТОЗОФОСФАТНЫЙ ПУТЬ ОКИСЛЕНИЯ УГЛЕВОДОВ
Открытие пути прямого окисления углеводов, или, как его называют, пентозофосфатного цикла, принадлежит О. Варбургу, Ф. Липману, Ф. Дикенсу и В.А. Энгельгарду. Расхождение путей окисления углеводов – классического (цикл трикарбоновых кислот, или цикл Кребса) и пентозофосфатного – начинается со стадии образования гексозомонофосфата. Если глюкозо-6-фосфат изомеризуется во фруктозо-6-фосфат, который фосфорилируется второй раз и превращается во фруктозо-1,6-бисфосфат, то в этом случае дальнейший распад углеводов происходит по обычному гликолитическому пути с образованием пировиноградной кислоты, которая, окисляясь до ацетил-КоА, затем «сгорает» в цикле Кребса.
Если второго фосфорилирования гексозо-6-монофосфата не происходит, то фосфорилированная глюкоза может подвергаться прямому окислению до фосфопентоз. В норме доля пентозофосфатного пути в количественном превращении глюкозы обычно невелика, варьирует у разных организмов и зависит от типа ткани и ее функционального состояния.
У млекопитающих активность пентозофосфатного цикла относительно высока в печени, надпочечниках, эмбриональной ткани и молочной железе в период лактации. Значение этого пути в обмене веществ велико. Он поставляет восстановленный НАДФН, необходимый для биосинтеза жирных кислот, холестерина и т.д. За счет пентозофосфатного цикла примерно на 50% покрывается потребность организма в НАДФН.
Другая функция пентозофосфатного цикла заключается в том, что он поставляет пентозофосфаты для синтеза нуклеиновых кислот и многих коферментов. При ряде патологических состояний удельный вес пенто-зофосфатного пути окисления глюкозы возрастает. Механизм реакций пентозофосфатного цикла достаточно расшифрован.
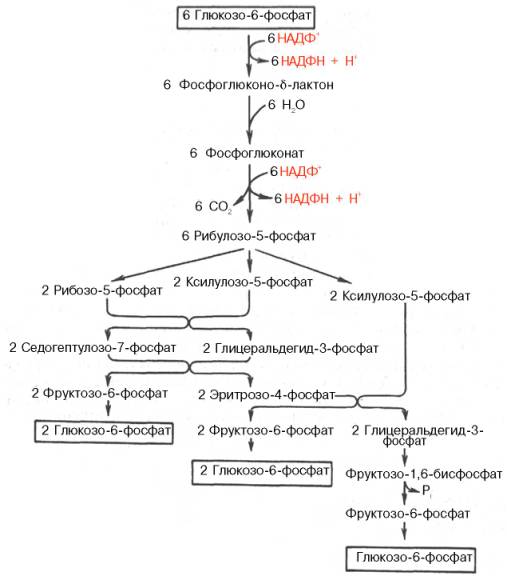
Рис. 10.12. Пентозофосфатный путь окисления углеводов.
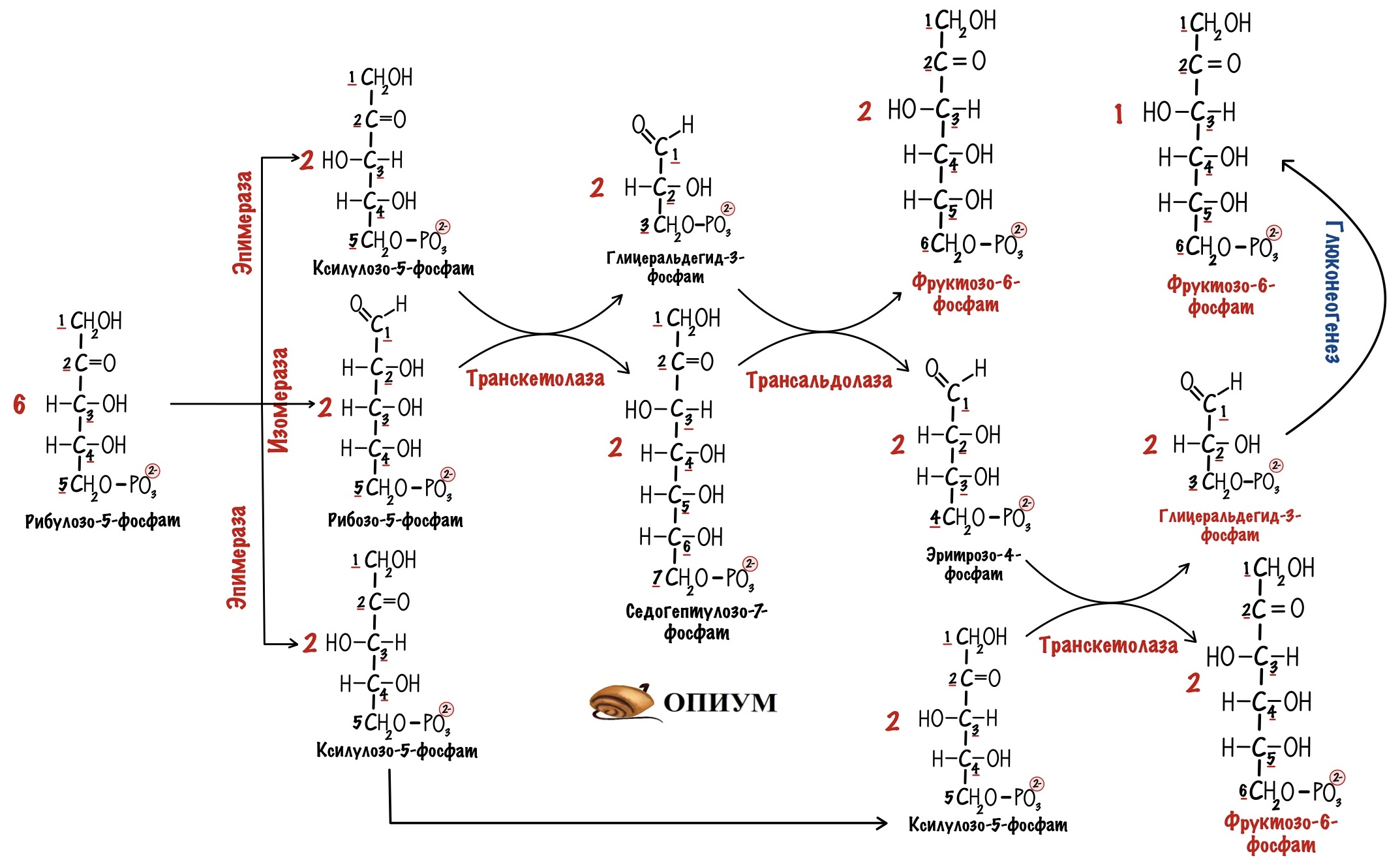
Пентозофосфатный цикл начинается с окисления глюкозо-6-фосфата и последующего окислительного декарбоксилирования продукта (в результате от гексозофосфата отщепляется первый атом углерода). Это первая, так называемая окислительная, стадия пентозофосфатного цикла. Вторая стадия включает неокислительные превращения пентозофосфатов с образованием исходного глюкозо-6-фосфата (рис. 10.12). Реакции пен-тозофосфатного цикла протекают в цитозоле клетки.
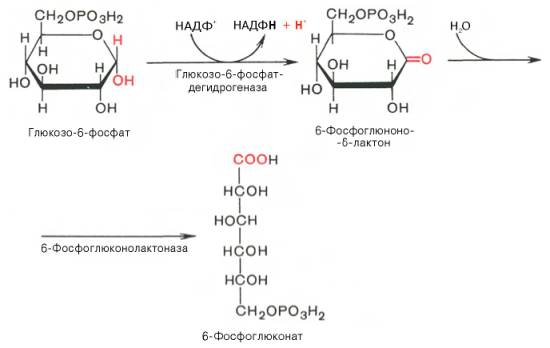
Первая реакция – дегидрирование глюкозо-6-фосфата при участии фермента глюкозо-6-фосфатдегидрогеназы и кофермента НАДФ + . Образовавшийся в ходе реакции 6-фосфоглюконо-δ-лактон – соединение нестабильное и с большой скоростью гидролизуется либо спонтанно, либо с помощью фермента 6-фосфоглюконолактоназы с образованием 6-фос-фоглюконовой кислоты (6-фосфоглюконат):
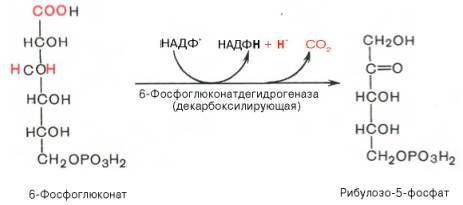
Во второй – окислительной – реакции, катализируемой 6-фосфоглюко-натдегидрогеназой (декарбоксилирующей), 6-фосфоглюконат дегидрируется и декарбоксилируется. В результате образуется фосфорилированная кетопентоза – D-рибулозо-5-фосфат и еще 1 молекула НАДФН:
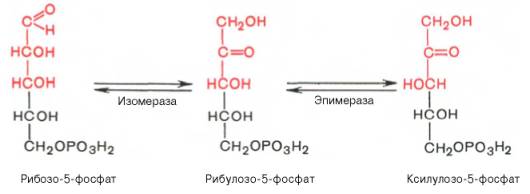
Под действием соответствующей эпимеразы из рибулозо-5-фосфата может образоваться другая фосфопентоза – ксилулозо-5-фосфат. Кроме того, рибулозо-5-фосфат под влиянием особой изомеразы легко превращается в рибозо-5-фосфат. Между этими формами пентозофосфатов устанавливается состояние подвижного равновесия:
При определенных условиях пентозофосфатный путь на этом этапе может быть завершен. Однако при других условиях наступает так называемый неокислительный этап (стадия) пентозофосфатного цикла. Реакции этого этапа не связаны с использованием кислорода и протекают в анаэробных условиях. При этом образуются вещества, характерные для первой стадии гликолиза (фруктозо-6-фосфат, фруктозо-1,6-бисфосфат, фосфотрио-зы), а другие – специфические для пентозофосфатного пути (седогептулозо-7-фосфат, пентозо-5-фосфаты, эритрозо-4-фосфат).
Основными реакциями неокислительной стадии пентозофосфатного цикла являются транскетолазная и трансальдолазная. Эти реакции катализируют превращение изомерных пентозо-5-фосфатов:
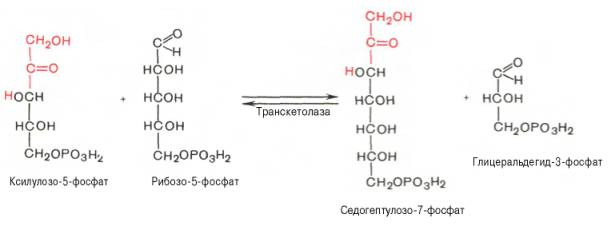
Коферментом в транскетолазной реакции служит ТПФ, играющий роль промежуточного переносчика гликольальдегидной группы от ксилулозо-5-фосфата к рибозо-5-фосфату. В результате образуется семиуглеродный моносахарид седогептулозо-7-фосфат и глицеральдегид-3-фосфат.
Транскетолазная реакция в пентозном цикле встречается дважды, второй раз – при образовании фруктозо-6-фосфата и триозофосфата в результате взаимодействия второй молекулы ксилулозо-5-фосфата с эритрозо-4-фосфатом:
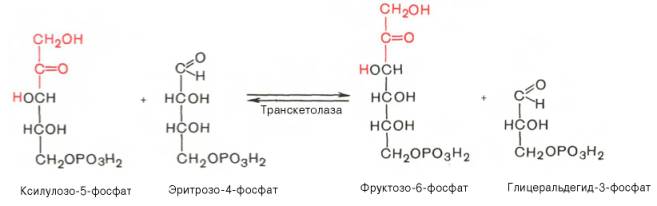
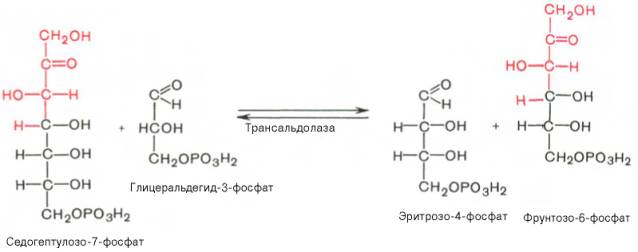
Фермент трансальдолаза катализирует перенос остатка диоксиацетона (но не свободного диоксиацетона) от седогептулозо-7-фосфата на гли-церальдегид-3-фосфат:
Шесть молекул глюкозо-6-фосфата, вступая в пентозофосфатный цикл, образуют 6 молекул рибулозо-5-фосфата и 6 молекул СО2, после чего из 6 молекул рибулозо-5-фосфата снова регенерируется 5 молекул глюко-зо-6-фосфата (см. рис. 10.12). Однако это не означает, что молекула глюкозо-6-фосфата, вступающая в цикл, полностью окисляется. Все 6 молекул СО2 образуются из С-1-атомов 6 молекул глюкозо-6-фосфата.
Валовое уравнение окислительной и неокислительной стадий пенто-зофосфатного цикла можно представить в следующем виде:

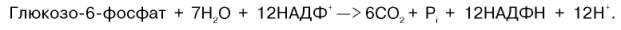
Образовавшийся НАДФН используется в цитозоле на восстановительные синтезы и, как правило, не участвует в окислительном фосфори-лировании, протекающем в митохондриях.
В последние годы появились работы, которые дают основание предполагать, что в некоторых тканях схема пентозофосфатного превращения углеводов сложнее, чем это представлено на рис. 10.12. Согласно этой более полной схеме пентозофосфатного пути, первые этапы превращения совпадают с прежней схемой, однако после первой транскетолазной реакции начинаются некоторые отклонения (рис. 10.13).
Считают, что пентозофосфатный путь и гликолиз, протекающие в ци-тозоле, взаимосвязаны и способны переключаться друг на друга в зависимости от соотношения концентраций промежуточных продуктов, образовавшихся в клетке (см. рис. 10.13).
БИОХИМИЯ УЧЕБНИК ДЛЯ ВУЗОВ — Е. С. Северина — 2004
РАЗДЕЛ 7. ОБМЕН УГЛЕВОДОВ
XII. Пентозофосфатный путь превращения глюкозы
Пентозофосфатный путь, называемый также гексомонофосфатным шунтом, служит альтернативным путём окисления глюкозо-6-фосфата. Пентозофосфатный путь состоит из 2 фаз (частей) — окислительной и неокислительной.
В окислительной фазе глюкозо-6-фосфат необратимо окисляется в пентозу — рибулозо-5- фосфат, и образуется восстановленный NADРН.
В неокислительной фазе рибулозо-5-фосфат обратимо превращается в рибозо-5-фосфат и метаболиты гликолиза.
Пентозофосфатный путь обеспечивает клетки рибозой для синтеза пуриновых и пиримидиновых нуклеотидов и гидрированным коферментом NАDРН, который используется в восстановительных процессах.
Суммарное уравнение пентозофосфатного пути выражается следующим образом:
3 Глюкозо-6-фосфат + 6 NАDР + —> 3 СO2 + 6 NADРН + Н + ) + 2 Фруктозо-6-фосфат + Глицеральдегид-3-фосфат.
Ферменты пентозофосфатного пути, так же, как и ферменты гликолиза, локализованы в цитозоле.
Наиболее активно пентозофосфатный путь протекает в жировой ткани, печени, коре надпочечников, эритроцитах, молочной железе в период лактации, семенниках.
А. Окислительный этап
В окислительной части пентозофосфатного пути глюкозо-6-фосфат подвергается окислительному декарбоксилированию, в результате которого образуются пентозы. Этот этап включает 2 реакции дегидрирования.
Первая реакция дегидрирования — превращение глюкозо-6-фосфата в глюконолактон-6- фосфат — катализируется NADP + -зaвиcимoй глюкозо-6-фосфатдегидрогеназой и сопровождается окислением альдегидной группы у первого атома углерода и образованием одной молекулы восстановленного кофермента NАDРH.
Далее глюконолактон-6-фосфат быстро превращается в 6-фосфоглюконат при участии фермента глюконолактонгидратазы.
Фермент 6-фосфоглюконатдегидрогеназа катализирует вторую реакцию дегидрирования окислительной части, в ходе которой происходит также и декарбоксилирование. При этом углеродная цепь укорачивается на один атом углерода, образуется рибулозо-5-фосфат и вторая молекула гидрированного NАDРН (рис. 7-62).
Рис. 7-62. Окислительный этап пентозофосфатного пути.

Восстановленный NАDРН ингибирует первый фермент окислительного этапа пентозофосфатного пути — глюкозо-6-фосфатдегидрогеназу. Превращение NАDРН в окисленное состояние NАDР + приводит к ослаблению ингибирования фермента. При этом скорость соответствующей реакции возрастает, и образуется большее количество NАDРН.
Суммарное уравнение окислительного этапа пентозофосфатного пути можно представить в виде:
Глюкозо-6-фосфат + 2 NАDР + + Н2O —> Рибулозо-5-фосфат + 2 (NАDРН + Н + ) + СO2.
Реакции окислительного этапа служат основным источником NАDРН в клетках. Гидрированные коферменты снабжают водородом биосинтетические процессы, окислительно-восстановительные реакции, включающие защиту клеток от активных форм кислорода. NАDРН как донор водорода участвует в анаболических процессах, например, в синтезе холестерина. Это источник восстановительных эквивалентов для цитохрома Р450, катализирующего образование гидроксильных групп при синтезе стероидных гормонов, жёлчных кислот, при катаболизме лекарственных веществ и других чужеродных соединений (см. разделы 8, 11, 12). Высокая активность фермента глюкозо-6- фосфатдегидрогеназы обнаружена в фагоцитирующих лейкоцитах, где NАDРН-оксидаза использует восстановленный NАDРН для образования супероксидного иона из молекулярного кислорода. Супероксидный ион генерирует другие активные формы кислорода, под действием которых и повреждаются молекулы ДНК, белков, липидов бактериальных клеток. Синтез жирных кислот из углеводов в печени является основным путём утилизации NADPH и обеспечивает регенерацию окисленной формы NADP + . В печени глюкозо-6- фосфатдегидрогеназа, как и ключевые ферменты гликолиза и биосинтеза жирных кислот, индуцируется при увеличении соотношения инсулин/глюкагон после приёма богатой углеводами пищи.
Несмотря на то, что NАDРН образуется также при окислении малата до пирувата и диоксида углерода (при участии NАDР + -зависимой малатдегидрогеназы) и дегидрировании изо- цитрата (при участии NADP + -зaвиcимoй изо- цитратдегидрогеназы), в большинстве случаев потребности клеток в восстановительных эквивалентах удовлетворяются за счёт пентозофосфатного пути.
Реакции окислительного пути протекают только в том случае, если восстановленный кофермент NАDРН возвращается в исходное окисленное состояние NАDР + при участии NADPH-зaвиcимыx дегидрогеназ (т. е. при условии использования гидрированного NАDРН в восстановительных процессах). Если потребности клетки в NADPH незначительны, рибозо-5-фосфат образуется в результате обратимых реакций неокислительного этапа пентозофосфатного пути, используя в качестве исходных веществ метаболиты гликолиза — глицеральдегид-3-фосфат и фруктозо-6-фосфат.
Б. Неокислительный этап
Неокислительный этап пентозофосфатного пути включает серию обратимых реакций, в результате которых рибулозо-5-фосфат превращается в рибозо-5-фосфат и ксилулозо-5-фосфат, и далее за счёт переноса углеродных фрагментов в метаболиты гликолиза — фруктозо-6-фосфат и глицеральдегид-3-фосфат. В этих превращениях принимают участие ферменты: эпимераза, изомераза, транскетолаза и трансальдолаза. Транскетолаза в качестве кофермента использует тиаминдифосфат. Неокислительный этап пентозофосфатного пути не включает реакции дегидрирования и поэтому используется только для синтеза пентоз.
Рибулозо-5-фосфат служит субстратом для двух ферментов. Фермент рибулозо-5-фосфат-3-эпимераза изменяет стехиометрическое положение одной ОН-группы у третьего атома углерода, превращая рибулозо-5-фосфат в ксилулозо-5-фосфат. Другой фермент — рибулозо-5-фосфат-изомераза — катализирует превращение рибулозо-5-фосфата в рибозо-5-фосфат (рис. 7-63). Рибозо-5- фосфат, образующийся в неокислительной фазе, обеспечивает клетки рибозой, необходимой для синтеза нуклеотидов, которые служат предшественниками и структурными компонентами коферментов дегидрогеназ и нуклеиновых кислот.
Рис. 7-63. Превращения рибулозо-5-фосфата.

Ферменты транскетолаза и трансальдолаза катализируют перенос двух- и трёхуглеродных фрагментов, соответственно используя в качестве донора углеродных фрагментов кетозу, а альдозу — в качестве акцептора. Эти реакции протекают в 2 этапа: сначала происходит отщепление углеродного фрагмента от молекулы-донора, а затем — перенос этого фрагмента на молекулу, выполняющую роль акцептора. Транскетолаза в неокислительной фазе пентозофосфатного пути катализирует 2 реакции. В первой реакции (рис. 7-64) транскетолаза расщепляет связь С-С между кетогруппой и соседним атомом углерода в молекуле ксилулозо-5-фосфат, в результате чего кетосахар превращается в альдозу, глицеральде- гид-3-фосфат, содержащую на 2 атома углерода меньше. Образующийся после расщепления двухуглеродный фрагмент остаётся ковалентно связанным в каталитическом центре фермента с коферментом тиаминдифосфатом. Далее фермент переносит двухуглеродный фрагмент на альдегидную группу альдосахара, образую новую кетозу — седогептулозо-7-фосфат.
Рис. 7-64. Реакция переноса двухуглеродного фрагмента, катализируемая транскетолазой.

Трансальдолаза переносит трёхуглеродный фрагмент от седогептулозо-7-фосфата на глицеральдегид-3-фосфат, образуя эритрозо-4-фосфат и фруктозо-6-фосфат (рис. 7-65).
Рис. 7-65. Реакция, катализируемая трансальдолазой.

Эта реакция подобна реакции альдольного расщепления гликолитического пути, за исключением того, что в данном случае трёхуглеродный фрагмент, содержащий кетогруппу, переносится на альдосахар глицеральдегид-3-фосфат, а в гликолитическом пути кетофрагмент высвобождается в виде дигидроксиацетонфосфата.
В следующей реакции, катализируемой транс- кетолазой, происходит перенос двухуглеродного фрагмента от ксилулозо-5-фосфата на эритрозо-4-фосфат. Продуктами этой реакции являются фруктозо-6-фосфат и глицеральдегид- 3-фосфат (рис. 7-66).
Рис. 7-66. Реакция, катализируемая транскетолазой.

Так как все реакции неокислительного этапа обратимы, образование рибозо-5-фосфата может происходить не только в результате изомерного превращения продукта окислительной фазы пентозофосфатного пути рибулозо-5-фосфата в рибозо-5-фосфат под действием изомеразы, но также и из промежуточных продуктов гликолиза — фруктозо-6-фосфата и глицеральдегид-3-фосфата. Последовательность превращений, приводящих к образованию рибозо-5- фосфата из таких продуктов гликолитического пути, можно представить в виде:
2 Фруктозо-6-фосфат + Глицералъдегид-3-фосфат —> 2 Ксилулозо-5-фосфат + Рибозо-5-фосфат
2 Ксилулозо-5-фосфат —> 2 Рибулозо-5-фосфат
2 Рибулозо-5-фосфат —> 2 Рибозо-5-фосфат.
Суммарный результат метаболизма 3 молекул рибулозо-5-фосфата в неокислительной фазе пентозофосфатного пути — образование 2 молекул фруктозо-6-фосфата и 1 молекулы глице- ральдегид-3-фосфата. Далее фруктозо-6-фосфат и глицеральдегид-3-фосфат могут превратиться в глюкозу. С учётом стехиометрического коэффициента, равного 2, для образования 5 молекул глюкозы (содержащих 30 атомов углерода) потребуются 4 молекулы фруктозо-6-фосфата и 2 молекулы глицеральдегид-3-фосфата (в сумме содержащие также 30 атомов углерода) или, соответственно, 6 молекул рибулозо-5-фосфата. Таким образом, неокислительный путь можно представить, как процесс возвращения пентоз в фонд гексоз.
В. Пентозофосфатный цикл
Окислительный этап образования пентоз и неокислительный этап (путь возвращения пентоз в гексозы) составляют вместе циклический процесс.
Такой процесс можно описать общим уравнением:
6 Глюкозо-6-фосфат + 12 NADР + + 2 Н2O —> 5 Глюкозо-6-фосфат + 12 NАDРН +12 Н + + 6 СO2.
Это означает, что из 6 молекул глюкозы образуются 6 молекул рибулозо-5-фосфат (пентозы) и 6 молекул СO2. Ферменты неокислительной фазы превращают 6 молекул рибулозо-5-фосфат в 5 молекул глюкозы (гексозы). При последовательном проведении этих реакций единственным полезным продуктом является NАDРН, образующийся в окислительной фазе пентозофосфатного пути. Такой процесс называют пентозофосфатным циклом (рис. 7-67).
Рис. 7-67. Пентозофосфатный цикл в жировой ткани.

Протекание пентозофосфатного цикла позволяет клеткам продуцировать NАDРН, необходимый для синтеза жиров, не накапливая пентозы.
Энергия, выделяющаяся при распаде глюкозы, трансформируется в энергию высокоэнергетического донора водорода — NАDРН. Гидрированный NАDРН служит источником водорода для восстановительных синтезов, а энергия NADPH преобразуется и сохраняется во вновь синтезированных веществах, например, жирных кислотах, высвобождается при их катаболизме и используется клетками.
Г. Дефект глюкозо-6-фосфат дегидрогеназы в эритроцитах
Неферментативное окисление гемоглобина (Fе 2+ ) в метгемоглобин (Fе 3+ ) приводит к одноэлектронному восстановлению кислорода и появлению реакционноспособного анион-радикала — супероксида O2-, который служит предшественником других активных форм кислорода: пероксида водорода Н2O2 и гидроксильного радикала ОН — . Активные формы кислорода являются сильнейшими окислителями и поэтому способны вызывать серьёзные повреждения молекул ДНК, белков, ненасыщенных липидов.
В эритроцитах, как и в большинстве клеток, присутствует тиолсодержащий трипептид — глутатион (y-глутамил-цистенил-глицин). Восстановленная форма глутатиона (Г-SН) содержит SН-группу (рис. 7-68), которая может служить донором электронов в реакциях восстановления. Под действием фермента глутатионпероксидазы восстановленный глутатион превращает молекулу пероксида водорода в молекулу воды, а сам переходит в окисленное состояние (Г-SS-Г). Регенерацию восстановленного глутатиона обеспечивает глутатионредуктаза, используя в качестве донора водорода гидрированный NАDРН. Для эритроцитов единственным источником получения NADРН служит пентозофосфатный путь, для других тканей существует альтернативный способ — при участии NАDН-зависимой малатдегидрогеназы (малик-фермент).
Рис. 7-68. Восстановление глутатиона под действием глутатионредуктазы. А — строение глутатиона; Б — восстановление глутатиона.

Взаимодействие восстановленного глутатиона: пероксидом водорода в эритроцитах предохраняет цистеиновые остатки в протомерах гемоглобина от окисления. При генетическом дефекте глюкозо-6-фосфатдегидрогеназы концентрация восстановленного кофермента NАDРН уменьшается, в результате чего резко снижается концентрация восстановленного глутатиона, а в клетке, соответственно, увеличивается количество активных форм кислорода. В этом случае окисление SН-групп молекул гемоглобина в эритроцитах приводит к образованию перекрёстных дисульфидных связей и агрегации протомеров гемоглобина с формированием телец Хайнца (см. раздел 14). В присутствии телец Хайнца пластичность мембраны нарушается, и она теряет способность к деформации при прохождении эритроцитов через капилляры. Это вызывает нарушение целостности мембраны, что приводит к гемолизу эритроцитов. Некоторые лекарственные вещества, например, антималярийный препарат примахин, сульфаниламиды, также снижают способность эритроцитов бороться с активными формами кислорода.
Биологическая библиотека — материалы для студентов, учителей, учеников и их родителей.
Наш сайт не претендует на авторство размещенных материалов. Мы только конвертируем в удобный формат материалы, которые находятся в открытом доступе и присланные нашими посетителями.
Если вы являетесь обладателем авторского права на любой размещенный у нас материал и намерены удалить его или получить ссылки на место коммерческого размещения материалов, обратитесь для согласования к администратору сайта.
Разрешается копировать материалы с обязательной гипертекстовой ссылкой на сайт, будьте благодарными мы затратили много усилий чтобы привести информацию в удобный вид.
© 2018-2022 Все права на дизайн сайта принадлежат С.Є.А.
Пентозофосфатный путь окисления глюкозы
Для начала нужно понять, что это за путь такой и какая молекула в него вступает. Пентозофосфатный путь — это путь окисления глюкозы. Точнее, не совсем глюкозы, а глюкозо-6-фосфата. Мы уже знаем, что глюкоза, которая попадает в клетку, фосфорилируется гексокиназой или глюкокиназой. В зависимости от типа ткани. В печени работает глюкокиназа, а в остальных тканях — гексокиназа. Клетка получает глюкозо-6-фосфат, который можно отправить на синтез гликогена или окислить до пирувата с помощью гликолиза.
Но существует другой путь окисления глюкозо-6-фосфата — пентозофосфатный. Если в ходе гликолиза клетка стремится получить энергию из этой молекулы, то в пентозофосфатном у неё другие цели. И вот какие: получить кофермент НАДФH и пентозы. И тут вы задаётесь вопросом: «Неужели эти молекулы так важны, что для них существует целый отдельный путь?». Да, важны — сейчас посмотрим почему.
Функции пентозофосфатного пути
Мы первый раз встречаемся с коферментом НАДФH, до этого речь шла только о НАДH. Этот восстановленный кофермент используется для некоторых химических реакций, но его основная роль — отдать два электрона в дыхательную цепь. В дыхательной цепи из электронов получится 2,5 молекулы АТФ. Но я отвлёкся.
НАД + отличается от НАДФ + всего на одну фосфатную группу, но при этом у них огромное различие в функциях.
Для чего же клетке нужен НАДФH?
- Синтеза жирных кислот;
- Синтеза холестерола;
- Синтеза нейромедиаторов;
- Образования дезоксирибозы — компонента нуклеотидов ДНК;
- Реакций обезвреживания ксенобиотиков и активных форм кислорода.
Довольно большой список. Даже если ты ещё не встречался с этими процессами, то он всё равно впечатляет. Видно, что у НАДФH огромная роль в нашем организме. И вот в чём прикол: пентозофосфатный путь почти единственный способ получить НАДФH для клеточек. Есть ещё малик-фермент и дегидрирование изоцитрата, но о них в другой раз. В норме клетке хватает пентозофосфатного пути для синтеза НАДФH.
Но это только первый продукт пентозофосфатного пути, а ведь были ещё пентозы. А если точнее, то в ходе пентозофосфатного пути образуется рибоза-5-фосфат. Она используется для синтеза:
- Нуклеотидов ДНК и РНК;
- АТФ;
- Коферментов НАД, НАДФ, ФАД и А.
У всех этих молекул есть кое-что общее. В их формулах есть рибоза, либо дезоксирибоза. А дезоксирибоза это производное чего? Рибозы. Респект.
Пентозофосфатный путь — поставщик НАДФH и пентоз для клетки.
Реакции пентозофосфатного пути
Реакции протекают в цитоплазме клетки, там же где и гликолиз. Пентозофосфатный путь состоит из двух этапов: окислительного и неокислительного. Мне не совсем нравится слово «этап». Оно подразумевает, что это два друг за другом следующих процесса. Но к концу статьи вы поймёте, что пентозофосфатный путь может идти не только последовательно, но и отдельно по каждому этапу. Ну вот, я опять забежал вперёд.
Окислительный этап
Мы договорились, что в пентозофосфатный путь вступает глюкозо-6-фосфат. В ходе окислительного этапа из этой молекулы образуется рибулозо-5-фосфат. Глюкоза теряет один атом углерода — из гексозы образуется пентоза. Давайте прикинем, что нужно для этого. Легко отщепить атом углерода, когда он карбоксильный — ведь у нас есть реакции декарбоксилирования. В них отщепляется кислотная группа COOH. В глюкозо-6-фосфате, увы, такой группы нет. Есть только альдегидная, да и та участвует в образовании циклической формы (вспомнить про альдозы и кетозы можно тут). Поэтому окислительный этап будет направлен на раскрытие цикла и перевод альдегидной группы в карбоксильную.
Первая реакция. Катализатор — глюкозо-6-фосфатдегидрогеназа. Кофермент реакции НАДФ. В ходе этой реакции от молекулы глюкозо-6-фосфата отщепляется два водорода. При этом НАДФ присоединяет к себе гидрид ион. Гидрид ион состоит из протона водорода и двух электронов. Образуется глюконолактон-6-фосфат и НАДФH. Один протон отправляется в цитоплазму.
Для того, чтобы превратить глюконолактон-6-фосфат в пентозу нужно раскрыть цикл. Ведь наша молекула до сих пор в циклической форме. За счёт дегидрирования мы увеличили нестабильность связи, с помощью которой образуется цикл. Теперь эту связь легко порвать.
Вторая реакция. Фермент — глюконолактон-гидратаза. У неё есть название попроще — лактоназа. Лактоназа присоединяет молекулу воды по нестабильной связи, что приводит к её разрушению. В ходе этой реакции образуется 6-фосфоглюконат.
Эти две реакции нужны были для раскрытия шестичленного цикла. Но не только. Ещё мы превратили альдегидную группу глюкозо-6-фосфата, которая находится у первого углерода, в карбоксильную группу. Или превратили альдегид в кислоту. Ведь реакция декарбоксилирования пойдёт только при наличии карбоксильной группы. Да-да, я капитан очевидность.
Третья реакция. Катализирует реакцию 6-фосфоглюконатдегидрогеназа, а её кофермент — НАДФ. В этой реакции происходят два важных момента: глюконат теряет карбоксильную группу, и от пентозы отщепляются два водорода. Продукты реакции — рибулозо-5-фосфат и НАДФH.
На этом заканчивается окислительный этап. Клетка синтезировала рибулозо-5-фосфат из глюкозо-6-фосфата. Она превратила шестиуглеродную альдозу в пятиуглеродную кетозу. И не менее важно, что клетка получила две молекулы НАДФH.
Итог окислительного этапа: образование пятиуглеродной кетозы и двух НАДФH
И ещё одно — все эти реакции были необратимыми. Этим окислительный этап отличается от неокислительного. Там все реакции будут обратимыми.
Неокислительный этап
Четвёртая реакция. Далее из рибулозо-5-фосфата можно получить рибозо-5-фосфат. Для этого нужно провести реакцию изомеризации. Ведь рибоза — это альдоза, а не кетоза. Фермент — фосфопентозоизомераза. Или просто изомераза.
Образуется рибозо-5-фосфат. Клетка получила то, что хотела — рибозо-5-фосфат. Теперь его можно пустить на синтез ДНК, РНК, АТФ, НАД, НАДФ, ФАД или кофермента А. Это в идеале.
Что, если клетке не нужны эти молекулы? Допустим, что сейчас ей всего достаточно — она укомплектована по полной. Но при этом ей нужен НАДФ. Клетка хочет синтезировать жирные кислоты или ей нужно что-то обезвредить. А двух молекул НАДФH как-то маловато. Что делать?
Можно сделать вот как. Пустим больше молекул глюкозо-6-фосфата на пентозофосфатный путь — получим больше НАДФH. Но при этом в клетке накапливаются пентозы, которые ей не нужны. Невыгодно. Глупо тратить глюкозо-6-фосфат, получая только половину плюшек.
Поэтому клетка делает по-другому. Она превращает рибозо-5-фосфат обратно в глюкозу. Но как? Ведь ты сказал, что окислительный этап необратим. А я и не врал, для этого нужен неокислительный этап. Вообще суть неокислительного этапа — перебрасывание углеродов с одной молекулы на другую. Сорри за спойлеры.
Вернёмся к третьей реакции. В ходе неё был получен рибулозо-5-фосфат. Давайте предположим, что мы пустили на пентозофосфатный путь три молекулы глюкозо-6-фосфата, а не одну. Не думай, почувствуй — дальше всё станет понятно. Клетка получит три молекулы рибулозо-5-фосфата. Из них одну мы превратим в рибозо-5-фосфат. А две другие в ксилулозо-5-фосфат. Их тоже будет два.
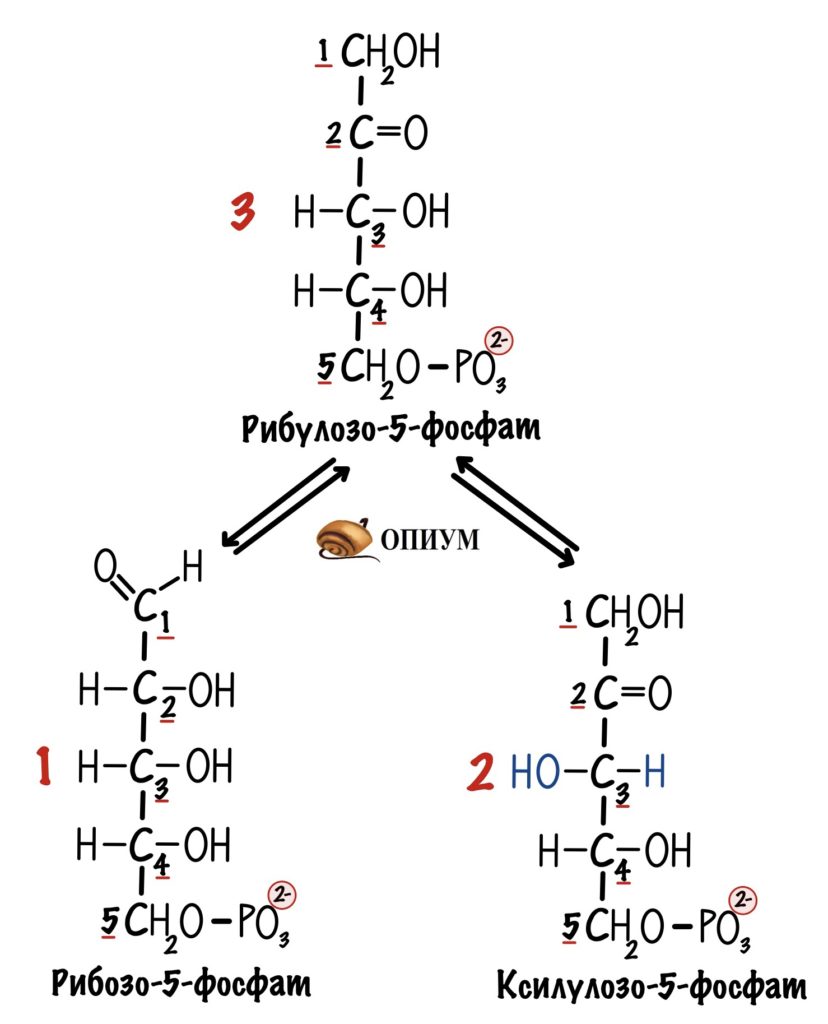
Фермент — фосфопентозоэпимераза. Или просто эпимераза. Она изменяет положение гидроксильной группы у третьего атома углерода. Теперь она слева, а не справа.
Можно начать синтезировать глюкозу из двух ксилулозо-5-фосфатов и одного рибозо-5-фосфата. Это невозможно сделать в одну реакцию, поэтому клетке понадобится целая серия. Сначала два атома углерода с ксилулозо-5-фосфата переносятся на рибозо-5-фосфат. Катализирует реакцию транскетолаза, она не может работать без своего кофермента — тиаминдифосфата.
В ходе этой реакции получаем глицеральдегид-3-фосфат и седогептулозо-7-фосфат (какое сложное название). Первый образовался из ксилулозо-5-фосфата. Для образования альдегидной группы нужна была реакция эпимеризации — превращения рибулозо-5-фосфата в ксилулозо-5-фосфат. А второй из рибозо-5-фосфата. Клетка получает триозу и гептозу.
Продолжаем перекидывать атомы углерода с одной молекулы на другую. Теперь у нас другой фермент — трансальдолаза. Она переносит три атома углерода с седогептулозо-7-фосфата на глицеральдегид-3-фосфат.
Образуется фруктозо-6-фосфат и эритрозо-4-фосфат. Вроде всё неплохо, фруктозо-6-фосфат можно превратить в глюкозу, но что делать с эритрозо-4-фосфатом? Вспоминаем, что у нас есть ещё одна молекула ксилулозо-5-фосфата. Снова используем транскетолазу. Она перекидывает два атома углерода с ксилулозо-5-фосфата на эритрозо-4-фосфат. Получаем ещё один фруктозо-6-фосфат и глицеральдегид-3-фосфат.
Получается, что из трёх молекул рибулозо-5-фосфата были синтезированы две молекулы фруктозо-6-фосфата и один глицеральдегид-3-фосфат. Из двух фруктозо-6-фосфатов образуется два глюкозо-6-фосфата.
А вот общая схема неокислительного этапа:
Глицеральдегид-3-фосфат — знакомая молекула, она встречалась нам в гликолизе. Это значит, что её можно отправить в глюконеогенез. Правда для того, чтобы синтезировать молекулу глюкозы нужно два глицеральдегид-3-фосфата. Ну давайте решим эту проблему тем, что удвоим количество глюкозо-6-фосфата, который был пущен на пентозофосфатный путь.
Мы взяли три молекулы, а теперь возьмём шесть. Из них будет синтезировано шесть молекул рибулозо-5-фосфата. Превратим их в две молекулы рибозо-5-фосфата и четыре молекулы ксилулозо-5-фосфата. Так мы получим четыре фруктозо-6-фосфата и два глицеральдегид-3-фосфата. Из фруктозо-6-фосфата образуется четыре молекулы глюкозо-6-фосфата, а две молекулы глицеральдегид-3-фосфата превратим в ещё один фруктозо-6-фосфат. Получается, что общее уравнение такого пентозофосфатного пути будет:
6 глюкозо-6-фосфат + 12НАДФ + + 6H2O = 5 глюкозо-6-фосфат + 12НАДФH + 12H + + 6CO2.
Поздравляю! Мы превратили наш пентозофосфатный путь в цикл. Пусть он и не будет работать слишком долго, но так мы уменьшим потери глюкозо-6-фосфата на образование НАДФH.
Пластичность пентозофосфатного пути
Ну а теперь посмотрим на пластичность пентозофосфатного пути в разных ситуациях.
Ситуация 1. Клетке нужны НАДФH и пентозы.
Допустим, что клетка готовится к делению. Ей нужно реплицировать ДНК и создать запас питательных веществ. На синтез ДНК нужны пентозы, а на запас — НАДФH.
Здесь всё просто — отправляем глюкозо-6-фосфат на окислительный этап. Далее превращаем рибулозо-5-фосфат в рибозо-5-фосфат. При этом получаем две молекулы НАДФH и одну пентозу. Использовать по потребности.
Ситуация 2. Клетке нужен НАДФH, но не нужны пентозы.
Такая ситуация будет в эритроцитах. В норме гемовое железо имеет заряд +2, но существуют метгемоглобинобразователи, которые переводят его в +3. При этом образуются активные формы кислорода. Им не место в эритроците — они могут повредить его мембрану. Повреждение мембраны приведёт к гемолизу. Вот мы и потеряли эритроцит. Но в эритроците есть герой — глутатион. Это трипептид, у него имеется -SH группа. Глутатион восстанавливает активные формы кислорода, отдавая электроны. А сам окисляется. Для того, чтобы вернуть глутатион в активное состояние нужен фермент глутатионпероксидаза. Она переносит электроны с НАДФH на глутатион, восстанавливая его. Вот такая сложная система.
Проводим окислительный этап, а далее зацикливаем процесс. Превращаем шесть молекул рибулозо-5-фосфата в пять молекул глюкозо-6-фосфата. Крутим, пока клетке не надоест этим заниматься. Эту ситуацию мы разбирали выше.
Ситуация 3. Клетке нужны пентозы, но не нужен НАДФH.
Допустим, что клетке требуются рибонуклеотиды, а дезоксирибонуклеотиды — нет. Значит ей нужен только рибозо-5-фосфат.
Забываем про окислительный этап. Помните, что в неокислительном этапе все реакции обратимы? Это значит, что мы можем повернуть их в другую сторону. В ходе гликолиза получаем четыре молекулы фруктозо-6-фосфата. Далее используем трансальдолазные и транскетолазные реакции для того, чтобы получить пять молекул рибозо-5-фосфата.
Правда для этого нам понадобится две молекулы глицеральдегид-3-фосфата, которые мы так же получим в ходе гликолиза. Но не трогая четыре фруктозо-6-фосфата, которые мы уже взяли до этого. Для этого пустим ещё одну молекулу глюкозо-6-фосфата в гликолиз.
Ситуация 4. Клетке нужны НАДФH и АТФ.
Такая ситуация тоже может быть в эритроцитах, когда им не нужно много НАДФH.
В этом случае будем отправлять глюкозо-6-фосфат на окислительный этап. Получаем пентозы и НАДФH. Пентозы с помощью транскетолазных и трансальдолазных реакций превращаем в фруктозо-6-фосфат и глицеральдегид-3-фосфат. Отправляем их в гликолиз. В эритроцитах идёт только анаэробный гликолиз, поэтому получаем лактат. В других клетках гликолиз будет аэробным — образуется пируват. Синтезированный пируват пойдёт в цикл трикарбоновых кислот.
Хочешь задать вопрос, похвалить или наговорить гадостей? Тогда залетай в телегу. Там ты сможешь предложить новый формат или разбор темы. А если серьёзно, то эти статьи пишутся для вас, поэтому мне важна обратная связь.
http://lifelib.info/biochemistry/biochemistry_4/49.html
http://opiumedu.ru/bihomiya/pentozofosfatnyj-put/