Контрольная Биохимия, вопросы 8, 15, 32, 46, 53,69 НГАУ
В работе вычислен pН буферного раствора, состоящий из 4 мл угольной кислоты и 16 мл гидрокарбоната натрия одинаковой концентрации, объяснен механизм образования электрического заряда на поверхности белковой молекулы, определено как заряжены частицы белка при рН = 9, если изоэлектрическая точка этого белка равна 8,5, написано уравнение реакции расщепления дипептидазой аланилцистеина, раскрыто что является конечным продуктом анаэробного окисления углеводов у млекопитающих животных, написано уравнение реакции образования молочной кислоты из пировиноградной, изучен внутриклеточный обмен липидов, составлено уравнение реакций образования тристеарина, написано уравнение реакции образования лецитина и кефалина. Учебное заведение — НГАУ
Авторство: Telesammit
Год: 2013 | Страниц: 10
8. Вычислить pН буферного раствора, состоящих из 4 мл угольной кислоты и 16 мл гидрокарбоната натрия одинаковой концентрации. Константа электрической диссоциации угольной кислоты 3,7∙10 -7
15. Объясните механизм образования электрического заряда на поверхности белковой молекулы. Как заряжены частицы белка при рН = 9, если изоэлектрическая точка этого белка равна 8,5?
32. Напишите уравнение реакции расщепления дипептидазой аланилцистеина.
46. Что является конечным продуктом анаэробного окисления углеводов у млекопитающих животных? Напишите уравнение реакции образования молочной кислоты из пировиноградной.
53. Внутриклеточный обмен липидов. Напишите уравнение реакций образования тристеарина.
69. Напишите уравнение реакции образования лецитина и кефалина.
Эта работа не подходит?
Если данная работа вам не подошла, вы можете заказать помощь у наших экспертов.
Оформите заказ и узнайте стоимость помощи по вашей работе в ближайшее время! Это бесплатно!
Эндопептидазы
Пепсин. Одним из хорошо изученных и основных протеолитических ферментов пищеварительного тракта является пепсин. Его наличие в желудке было установлено еще в 1783 г. Л. Спалланцани, хотя в кристаллическом виде он был получен только в 1930 г. (см. главу 1). Пепсин вырабатывается в главных клетках слизистой оболочки желудка в неактивной форме – в виде пепсиногена. Превращение пепсиногена в активный пепсин происходит в желудочном содержимом, однако молекулярный механизм этого превращения в деталях еще не выяснен. Наиболее вероятным считается предположение, что этот процесс является последовательным и протекает в несколько этапов в присутствии соляной кислоты по механизму аутокаталитического действия самого пепсина. Молекулярная масса пеп-синогена составляет приблизительно 40400, а пепсина – 32700, поэтому превращение первого во второй связано с отщеплением пептидных фрагментов. Оба фермента можно сравнительно легко получить в кристаллическом виде. Следует отметить, что в отличие от других протеиназ пепсин отличается высокой устойчивостью в сильнокислой среде и характеризуется низким значением изоэлектрической точки (рI 2+ .
Активирование трипсиногена химически выражается в отщеплении с N-конца полипептидной цепи 6 аминокислотных остатков (Вал–Асп– Асп–Асп–Асп–Лиз) и соответственно в укорочении полипептидной цепи (рис. 12.1).
Следует подчеркнуть, что в этом небольшом, казалось бы, химическом процессе – отщепление гексапептида от предшественника – заключено важное биологическое значение, поскольку при этом происходят формирование активного центра и образование трехмерной структуры трипсина, а известно (см. главы 1 и 4), что и белки биологически активны только в своей нативной трехмерной конформации. В том, что трипсин, как и другие протеиназы, вырабатывается в поджелудочной железе в неактивной форме, также имеется определенный физиологический смысл, поскольку в противном случае трипсин мог бы оказывать разрушающее протеолитическое действие не только на клетки самой железы, но и на другие ферменты, синтезируемые в ней (амилаза, липаза и др.). В то же время поджелудочная железа защищает себя еще одним механизмом – синтезом специфического белка ингибитора панкреатического трипсина. Этот ингибитор оказался
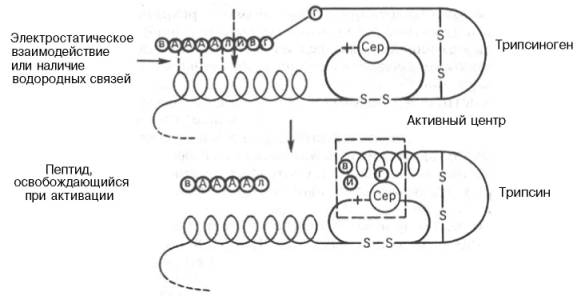
Рис. 12.1. Механизм активации трипсиногена быка (схема).
низкомолекулярным пептидом (мол. масса 6000), который прочно связывается с активными центрами трипсина и химотрипсина, вызывая обратимое их ингибирование. В поджелудочной железе синтезируется также α1-антипротеиназа (мол. масса 50000), которая преимущественно инги-бирует эластазу.
При остром панкреатите, когда трипсин и другие ферменты из пораженной поджелудочной железы «вымываются» в кровь, уровень их в крови соответствует размерам некротического участка. В этом случае определение активности трипсина в сыворотке крови является надежным ферментным тестом при диагностике острого панкреатита. Следует отметить, что субстратная специфичность трипсина ограничена разрывом только тех пептидных связей, в образовании которых участвуют карбоксильные группы лизина и аргинина.
Химотрипсин. В поджелудочной железе синтезируется ряд химотрип-синов (α-, β- и π-химотрипсины) из двух предшественников – химотрипси-ногена А и химотрипсиногена В. Активируются проферменты в кишечнике под действием активного трипсина и химотрипсина. Полностью раскрыта последовательность аминокислот химотрипсиногена А, во многом сходная с последовательностью аминокислот трипсина. Молекулярная масса его составляет примерно 25000. Он состоит из одной полипептидной цепи, содержащей 246 аминокислотных остатков. Активация профермента не сопряжена с отщеплением большого участка молекулы (см. рис. 4.3). Получены доказательства, что разрыв одной пептидной связи между аргинином и изолейцином в молекуле химотрипсиногена А под действием трипсина приводит к формированию π-химотрипсина, обладающего наибольшей ферментативной активностью. Последующее отщепление дипеп-тида Сер–Арг приводит к образованию δ-химотрипсина. Аутокаталити-ческий процесс активирования, вызванный химотрипсином, сначала способствует формированию неактивного промежуточного неохимотрипсина, который под действием активного трипсина превращается в α-химотрип-син; этот же продукт образуется из δ-химотрипсина, но под действием активного химотрипсина.
Таким образом, благодаря совместному перекрестному воздействию химотрипсина и трипсина из химотрипсиногена образуются разные химо-трипсины, различающиеся как ферментативной активностью, так и некоторыми физико-химическими свойствами, в частности электрофорети-ческой подвижностью.
Следует отметить, что химотрипсин обладает более широкой субстратной специфичностью, чем трипсин. Он катализирует гидролиз не только пептидов, но и эфиров, гидроксаматов, амидов и других ацилпроизводных, хотя наибольшую активность химотрипсин проявляет по отношению к пептидным связям, в образовании которых принимают участие карбоксильные группы ароматических аминокислот: фенилаланина, тирозина и триптофана.
Эластаза. В поджелудочной железе синтезируется еще одна эндопеп-тидаза – эластаза – в виде проэластазы. Превращение профермента в элас-тазу в тонкой кишке катализируется трипсином. Название фермент получил от субстрата эластина, который он гидролизует. Эластин содержится в соединительной ткани и характеризуется наличием большого числа остатков глицина и серина. Эластаза обладает широкой субстратной специфичностью, но предпочтительнее гидролизует пептидные связи, образованные аминокислотами с небольшими гидрофобными радикалами, в частности глицином, аланином и серином. Интересно, что ни трипсин, ни химотрипсин не гидролизуют пептидные связи молекулы эластина, хотя все три фермента, включая эластазу, содержат сходные участки аминокислотных последовательностей и одинаковые места положения дисульфидных мостиков, а также имеют в активном центре один и тот же ключевой остаток серина (см. табл. 4.2), что подтверждают опыты с ингибированием всех трех ферментов диизопропилфторфосфатом, химически связывающим ОН-группу серина. Высказано предположение, что все три эндопептидазы поджелудочной железы: трипсин, химотрипсин и эластаза,– возможно, имеют один и тот же общий предшественник и что специфичность активного фермента в основном определяется конформационными изменениями профермента в процессе активирования.
Экзопептидазы. В переваривании белков в тонкой кишке активное участие принимает семейство экзопептидаз. Одни из них – карбоксипеп-тидазы – синтезируются в поджелудочной железе в виде прокарбоксипеп-тидазы и активируются трипсином в кишечнике; другие – аминопептидазы – секретируются в клетках слизистой оболочки кишечника и также активируются трипсином.
Карбоксипептидазы. Подробно изучены две карбоксипептидазы – А и В, относящиеся к металлопротеинам и катализирующие отщепление от полипептида С-концевых аминокислот. Карбоксипептидаза А разрывает преимущественно пептидные связи, образованные концевыми ароматическими аминокислотами, а карбоксипептидаза В – связи, в образовании которых участвуют С-концевые лизин и аргинин. Очищенный препарат карбокси-пептидазы А обладает бифункциональной активностью – пептидазной и эстеразной и содержит ион Zn 2+ (один атом на 1 моль фермента). При замене ионов Zn 2+ на ионы Са 2+ полностью утрачивается пепти-дазная активность, но усиливается исходная эстеразная активность, хотя
при этом существенных изменений в третичной структуре фермента не отмечается.
Аминопептидазы. В кишечном соке открыты два фермента – аланин-аминопептидаза, катализирующая преимущественно гидролиз пептидной связи, в образовании которой участвует N-концевой аланин, и лейцин-аминопептидаза, не обладающая строгой субстратной специфичностью и гидролизующая пептидные связи, образованные любой N-концевой аминокислотой. Оба фермента осуществляют ступенчатое отщепление аминокислот от N-конца полипептидной цепи.
Дипептидазы. Процесс переваривания пептидов, их расщепление до свободных аминокислот в тонкой кишке завершают дипептидазы. Среди дипептидаз кишечного сока хорошо изучена глицилглицин-дипептидаза, гидролизующая соответствующий дипептид до двух молекул глицина. Известны также две другие дипептидазы: пролил-дипептидаза (пролиназа), катализирующая гидролиз пептидной связи, в образовании которой участвует СООН-группа пролина, и пролин-дипептидаза (пролидаза), гидроли-зующая дипептиды, в которых азот пролина связан кислотно-амидной связью.
Еще сравнительно недавно протеиназы традиционно связывали только с процессами переваривания. В настоящее время появляется все больше данных о более широкой биологической роли протеолитических ферментов органов и тканей в регуляции ряда вне- и внутриклеточных процессов. Некоторые протеиназы выполняют защитную функцию (свертывание крови, система комплемента, лизис клеток), другие генерируют гормоны, токсины, вазоактивные агенты (ангиотензин, кинины). Ряд протеиназ регулирует образование пищеварительных ферментов, взаимодействие между клетками и клеточными поверхностями, процессы фертилизации (хитин-синтетаза) и дифференциации. Регуляция в большинстве случаев предусматривает превращение неактивного предшественника в активный белок путем отщепления ограниченного числа пептидов. Этот процесс, впервые описанный К. Линдерстрем-Лангом еще в 50-е годы, в последнее время называют ограниченным протеолизом. Значение его очень важно для понимания сущности биологического синтеза в клетках неактивных пре-и пробелков. Кроме того, этот процесс нашел широкое практическое применение в лабораториях и промышленности. В регуляции действия протеолитических ферментов участвуют также ингибиторы протеиназ белковой природы, открытые не только в поджелудочной железе, но и в плазме крови, курином яйце и т.д.
Отделение панкреатического и кишечного соков регулируется нейро-гормональными факторами, которые подробно излагаются в курсе физиологии. Имеются доказательства роли соляной кислоты в качестве пускового механизма выработки в кишечнике особых гормонов. В частности, соляная кислота, попадая в двенадцатиперстную кишку, стимулирует секрецию секретина (см. главу 8); последний, стимулируя секрецию и отделение щелочного панкреатического сока, способствует оттоку желчи. Показано, что секретин быстро исчезает из кровотока, а новые порции его не вырабатываются, поскольку соляная кислота нейтрализуется щелочным панкреатическим соком. Таким образом, благодаря существованию такого механизма, действующего по типу обратной связи, осуществляется регуляция секреции и отделения поджелудочного сока. Поджелудочный сок, полученный при действии секретина, содержит незначительное количество ферментов, но богат бикарбонатами, создающими слабощелочную среду (рН 7,5–8,5), оптимальную для действия пищеварительных ферментов
в кишечнике. Вторым гормоном, также синтезирующимся в двенадцатиперстной кишке и регулирующим секрецию поджелудочного сока, является холецистокинин (панкреозимин); он стимулирует отделение сока, богатого ферментами и бедного бикарбонатами.
Acetyl
Наведите курсор на ячейку элемента, чтобы получить его краткое описание.
Чтобы получить подробное описание элемента, кликните по его названию.
| H + | Li + | K + | Na + | NH4 + | Ba 2+ | Ca 2+ | Mg 2+ | Sr 2+ | Al 3+ | Cr 3+ | Fe 2+ | Fe 3+ | Ni 2+ | Co 2+ | Mn 2+ | Zn 2+ | Ag + | Hg 2+ | Pb 2+ | Sn 2+ | Cu 2+ | |
| OH — | Р | Р | Р | Р | Р | М | Н | М | Н | Н | Н | Н | Н | Н | Н | Н | — | — | Н | Н | Н | |
| F — | Р | М | Р | Р | Р | М | Н | Н | М | М | Н | Н | Н | Р | Р | Р | Р | Р | — | Н | Р | Р |
| Cl — | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Н | Р | М | Р | Р |
| Br — | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Н | М | М | Р | Р |
| I — | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | ? | Р | ? | Р | Р | Р | Р | Н | Н | Н | М | ? |
| S 2- | М | Р | Р | Р | Р | — | — | — | Н | — | — | Н | — | Н | Н | Н | Н | Н | Н | Н | Н | Н |
| HS — | Р | Р | Р | Р | Р | Р | Р | Р | Р | ? | ? | ? | ? | ? | Н | ? | ? | ? | ? | ? | ? | ? |
| SO3 2- | Р | Р | Р | Р | Р | Н | Н | М | Н | ? | — | Н | ? | Н | Н | ? | М | М | — | Н | ? | ? |
| HSO3 — | Р | ? | Р | Р | Р | Р | Р | Р | Р | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? |
| SO4 2- | Р | Р | Р | Р | Р | Н | М | Р | Н | Р | Р | Р | Р | Р | Р | Р | Р | М | — | Н | Р | Р |
| HSO4 — | Р | Р | Р | Р | Р | Р | Р | Р | — | ? | ? | ? | ? | ? | ? | ? | ? | ? | ? | Н | ? | ? |
| NO3 — | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | Р | — | Р |
| NO2 — | Р | Р | Р | Р | Р | Р | Р | Р | Р | ? | ? | ? | ? | Р | М | ? | ? | М | ? | ? | ? | ? |
| PO4 3- | Р | Н | Р | Р | — | Н | Н | Н | Н | Н | Н | Н | Н | Н | Н | Н | Н | Н | Н | Н | Н | Н |
| CO3 2- | Р | Р | Р | Р | Р | Н | Н | Н | Н | ? | ? | Н | ? | Н | Н | Н | Н | Н | ? | Н | ? | Н |
| CH3COO — | Р | Р | Р | Р | Р | Р | Р | Р | Р | — | Р | Р | — | Р | Р | Р | Р | Р | Р | Р | — | Р |
| SiO3 2- | Н | Н | Р | Р | ? | Н | Н | Н | Н | ? | ? | Н | ? | ? | ? | Н | Н | ? | ? | Н | ? | ? |
| Растворимые (>1%) | Нерастворимые ( Спасибо! Ваша заявка отправлена, преподаватель свяжется с вами в ближайшее время. Вы можете также связаться с преподавателем напрямую: 8(906)72 3-11-5 2
Скопируйте эту ссылку, чтобы разместить результат запроса » » на другом сайте. Изображение вещества/реакции можно сохранить или скопировать, кликнув по нему правой кнопкой мыши. Если вы считаете, что результат запроса » » содержит ошибку, нажмите на кнопку «Отправить». Этим вы поможете сделать сайт лучше. К сожалению, регистрация на сайте пока недоступна. На сайте есть сноски двух типов: Подсказки — помогают вспомнить определения терминов или поясняют информацию, которая может быть сложна для начинающего. Дополнительная информация — такие сноски содержат примечания или уточнения, выходящие за рамки базовой школьной химии, нужны для углубленного изучения. Здесь вы можете выбрать параметры отображения органических соединений. источники: http://xumuk.ru/biologhim/181.html http://acetyl.ru/f/r184.php |