Математическая биология
Курс лекций «Математические модели в биологии»
 читается автором для студентов 2-ого года обучения бакалавриата Биологического факультета МГУ имени М. В. Ломоносова. Параллельно с лекциями проходят семинары (практические занятия), в ходе которых студенты закрепляют полученные на лекциях знания и знакомятся с программным обеспечением, используемым для анализа математических моделей и проведения вычислительных экспериментов. После прохождения курса студенты сдают экзамен. Курс включает 16 лекций по 2 академических часа.
читается автором для студентов 2-ого года обучения бакалавриата Биологического факультета МГУ имени М. В. Ломоносова. Параллельно с лекциями проходят семинары (практические занятия), в ходе которых студенты закрепляют полученные на лекциях знания и знакомятся с программным обеспечением, используемым для анализа математических моделей и проведения вычислительных экспериментов. После прохождения курса студенты сдают экзамен. Курс включает 16 лекций по 2 академических часа.
- Учебник Ризниченко Г. Ю.Лекции по математическим моделям в биологии (изд. 2-е, испр. и дополн.) Издательство РХД, 2011 г. 560 стр. ISBN 978-5-93972-847-8. Предыдущее издание (значительно более краткое!) находится в свободном доступе в сети Интернет по ссылке http://www.library.biophys.msu.ru/LectMB/
- Учебник Мятлев В.Д., Панченко Л.А., Ризниченко Г.Ю., Терёхин А.Т. Теория вероятностей и математическая статистика. Математические модели (изд. 2-е, испр. и дополн.) М.: Издательство Юрайт, 2018. — 321 с. — (Серия : Университеты России). — ISBN 978-5-534-01698-7.
- Учебное пособие Плюснина Т.Ю., Фурсова П. В., Дьяконова А. Н., Тёрлова Л. Д., Ризниченко Г. Ю.Математические модели в биологии (Изд. 3-e доп. Учебное пособие. М.-Ижевск: НИЦ: «Регулярная и хаотическая динамика», 2021. 174 с. ISBN: 978-5-4344-0922-3) — электронная версия
Лекции будут прочитаны в дистанционном режиме по четвергам с сентября по декабрь 2021 года.
2 сентября. Лекция 1. Вводная. Одно уравнение. Устойчивость
Часть 1. Введение. Понятие модели. Объекты, цели и методы моделирования. Модели в разных науках. Компьютерные и математические модели. История первых моделей в биологии. Современная классификация моделей биологических процессов. Регрессионные, имитационные, качественные модели. Принципы имитационного моделирования и примеры моделей. Специфика моделирования живых систем.
Часть 2. Модели биологических систем, описываемые одним дифференциальным уравнением первого порядка. Модели, приводящие к одному дифференциальному уравнению. Понятие решения одного автономного дифференциального уравнения. Стационарное состояние (состояние равновесия). Устойчивость состояния равновесия. Методы оценки устойчивости.
9 сентября. Лекция 2. Модели роста популяций (1)
Непрерывные модели: экспоненциальный рост, логистический рост, модели с наименьшей критической численностью. Модель роста человечества. Модели с неперекрывающимися поколениями. Дискретное логистическое уравнение. Диаграмма и лестница Ламерея. Типы решений при разных значениях параметра: монотонные и затухающие решения, циклы, квазистохастическое поведение, вспышки численности. Матричные модели популяций. Влияние запаздывания. Вероятностные модели популяций.
16 сентября Лекция 3. Модели роста популяций (2). Модели, описываемые системами двух автономных дифференциальных уравнений
Часть 2. Модели, описываемые системами двух автономных дифференциальных уравнений. Фазовая плоскость. Фазовый портрет. Метод изоклин. Главные изоклины. Устойчивость стационарного состояния. Линейные системы. Типы особых точек: узел, седло, фокус, центр. Метод Ляпунова линеаризации систем в окрестности стационарного состояния. Метод функции Ляпунова.
23 сентября Лекция 4. Базовые модели математической биологии, представленные двумя автономными уравнениями
Примеры исследования устойчивости стационарных состояний моделей биологических систем. Химические реакции первого порядка. Уравнения Лотки. Уравнения Вольтерра.
30 сентября Лекция 5. Мультистационарные системы. Типы бифуркаций. Катастрофы
Триггер. Примеры систем с двумя устойчивыми стационарными состояниями. Силовое и параметрическое переключение триггера. Эволюция. Отбор одного из двух и нескольких равноправных видов. Конкуренция двух видов в случае неограниченного и ограниченного роста. Генетический триггер Жакоба и Моно. Бифуркации динамических систем. Типы бифуркаций. Бифуркационные диаграммы и фазопараметрические портреты. Катастрофы.
- Программа: Мультистационарные системы
- Учебник: Мультистационарные системы
- Учебник: Проблема быстрых и медленных переменных. Теорема Тихонова. Типы бифуркаций. Катастрофы
- Материалы по теории катастроф:
- Арнольд В.И.Теория катастроф // Наука и жизнь, 1989, № 10
- Арнольд В.И.Теория катастроф // Динамические системы – 5, Итоги науки и техн. Сер. Соврем. пробл. мат. Фундам. направления, 5, ВИНИТИ, М., 1986, 219–277
- Арнольд В.И.Теория катастроф. М., Наука, 1990 — 128 с.
- Просмотр лекции в YouTube
- Загрузить видео (MP4)
- Демонстрация PowerPoint с голосовым сопровождением
7 октября Лекция 6. Проблема быстрых и медленных переменных. Теорема Тихонова
Метод квазистационарных концентраций. Теорема Тихонова. Уравнение Михаэлиса-Ментен. Формула Моно. Конкуренция двух видов, питающихся одинаковым субстратом.
14 октября Лекция 7. Колебания в биологических системах
Понятие автоколебаний. Изображение поведения автоколебательной системы на фазовой плоскости. Предельные циклы. Условия существования предельных циклов. Рождение предельного цикла. Бифуркация Андронова-Хопфа. Мягкое и жесткое возбуждение колебаний. Модель Брюсселятор. Примеры автоколебательных моделей процессов в живых системах. Колебания в темновых процессах фотосинтеза. Автоколебания в модели гликолиза. Внутриклеточные колебания концентрации кальция.
21 октября Лекция 8. Динамический хаос. Модели биологических сообществ. Фракталы
Основные понятия теории динамических систем. Предельные множества. Аттракторы. Странные аттракторы. Динамический хаос. Линейный анализ устойчивости траекторий. Диссипативные системы. Устойчивость хаотических решений. Размерность странных аттракторов. Стационарные состояния и динамические режимы в сообществе из трех видов. Динамический хаос в моделях взаимодействия видов. Трофические системы с фиксированным количеством вещества. Модель системы четырех биологических видов.
Фракталы и фрактальная размерность. Кривая Коха. Треугольник и салфетка Серпинского. Канторово множество. Канторов стержень, чертова лестница. Примеры фрактальных множеств в живых системах. Формирование крон деревьев. Альвеолы легких. Мембраны митохондрий.
28 октября Лекция 9. Модели взаимодействия видов
Гипотезы Вольтерра. Аналогии с химической кинетикой. Вольтерровские модели взаимодействий. Классификация типов взаимодействий. Конкуренция. Хищник-жертва. Обобщенные модели взаимодействия видов. Модель Колмогорова. Модель взаимодействия двух видов насекомых МакАртура. Пространственно-временные агентные модели взаимодействия видов.
11 ноября Лекция 10. Моделирование микробных популяций. Модели эпидемий
Часть 1. Моделирование микробных популяций. Микробные популяции как объект моделирования и управления. Непрерывная культура микроорганизмов. Модель Моно. Микроэволюционные процессы в микробных популяциях. Возрастные распределения. Двухвозрастная модель. Непрерывные возрастные распределения.
Часть 2. Модели эпидемий. Эпидемия в замкнутой популяции. Эпидемиологическая кривая. Классическая модель SIR. Базовое репродуктивное число инфекции. Модель COVID-19.
- Просмотр презентации в YouTube
- Загрузить видео презентации (MP4)
- Демонстрация PowerPoint с голосовым сопровождением
18 ноября Лекция 11. Распределенные биологические системы
Уравнение реакция-диффузия. Почему возникают периодические структуры и волны. ААктивные кинетические среды в живых системах. Проблема формообразования. Распространение волн возбуждения. Пространственные структуры и автоволновые процессы в химических и биохимических реакциях.
Уравнение диффузии. Начальные и граничные условия. Решение уравнения диффузии. Решение однородного уравнения диффузии с нулевыми граничными условиями. Метод разделения переменных. Собственные значения и собственные функции задачи Штурма-Лиувилля. Решение неоднородного уравнения с нулевыми начальными условиями. Решение общей краевой задачи. Линейный анализ устойчивости гомогенных стационарных решений одного уравнения типа реакция-диффузия.
Устойчивость однородных стационарных решений системы двух уравнений типа реакция-диффузия. Диссипативные структуры. Линейный анализ устойчивости гомогенного стационарного состояния. Зависимость вида неустойчивости от волнового числа. Неустойчивость Тьюринга. Линейный анализ устойчивости гомогенного стационарного состояния распределенного Брюсселятора. Диссипативные структуры вблизи порога неустойчивости. Локализованные диссипативные структуры. Линейный анализ системы реакция-электродиффузия. Типы пространственно-временных режимов.
25 ноября Лекция 12. Распределенные триггеры и морфогенез. Модели раскраски шкур животных
Распределенные триггеры и морфогенез. Модели раскраски шкур животных. Дифференциация и морфогенез. Модель генетического триггера с диффузией (Чернавский и др.). Исследование устойчивости гомогенного стационарного состояния. Генетический триггер с учетом диффузии субстратов. Модель гидры Гирера-Майнхардта. Моделирование раскраски шкур животных. Модели агрегации амеб.
30 ноября Контрольная работа
2 декабря Лекция 13. Распространение импульсов, фронтов и волн. Модели распространения нервного импульса. Автоволновые процессы и сердечные аритмии
Распространение импульсов, фронтов и волн. Модель распространения фронта волны Петровского-Колмогорова-Пискунова-Фишера. Взаимодействие процессов размножения и диффузии. Локальные функции размножения. Автомодельная переменная. Распространение амброзиевого листоеда.
Модели распространения нервного импульса. Автоволновые процессы и сердечные аритмии. Распространение нервного импульса. Опыты и модель Ходчкина-Хаксли. Редуцированная модель ФитцХью-Нагумо. Возбудимый элемент локальной системы. Подпороговое и надпороговое возбуждение. Бегущие импульсы. Детальные модели кардиоцитов. Аксиоматические модели возбудимой среды. Автоволновые процессы и сердечные аритмии.
Реакция Белоусова-Жаботинского – базовая модель нелинейного пространственно-временного поведения. Модель образования зон кислотного и щелочного рН вдоль мембраны клеточной водоросли Chara corallina.
9 декабря Лекция 14. Молекулярное моделирование
Иерархия масштабов биологических систем и типы моделей. Квантово-механические методы. Основы метода молекулярной динамики. Рентгеноструктурные данные. Библиотеки фрагментов. Генерация трехмерных координат. Потенциалы молекулярных взаимодействий. Совмещение молекулярных полей. Принципы организации структуры белков. Моделирование белков по гомологии. Процедуры оптимизации. Валидация моделей белков. Виртуальный скрининг и докинг. Разработка лекарственных веществ с использованием методов молекулярного моделирования. Компьютерные пакеты.
16 декабря Лекция 15. Кинетические и Монте Карло модели процессов в фотосинтетической мембране
Модели биологического электронного транспорта. Окислительно-восстановительные реакции в растворе и в мультиферментных комплексах, локализованных в мембране. Процессы в энергопреобразующих мембранах митохондрий и хлоропластов. Перенос электрона в мультиферментном комплексе. Графы состояний. Примеры переноса электрона в комплексах двух и трех переносчиков. Обмен электронами комплекса с подвижными переносчиками. Модель переноса электронов в изолированной фотосистеме 2. Упрощенная и детальная кинетические модели взаимодействия фотосистем 1 и 2. Метод Монте-Карло моделирования процессов в ансамблях фотосинтетических цепей.
23 декабря Лекция 16. Броуновские и молекулярные модели первичных процессов фотосинтеза
Компьютерное многочастичное моделирование. Броуновское движение подвижных переносчиков и их электростатические взаимодействия с мембранными комплексами. Броуновское описание образования предварительного комплекса белков – донора и акцептора электрона. Молекулярное описание процессов образования финального комплекса. Роль сложного интерьера клетки в кинетике наблюдаемых процессов. Модели, использующие разные типы описания процессов в фотосинтетической мембране: электронный транспорт внутри комплексов, взаимодействие подвижных белков-переносчиков с фотосинтетическим реакционным центром, пространственно-временная эволюция протонного потенциала.
Основные понятия моделей описываемых одним уравнением
Г.Ю. Ризниченко — профессор, био-фак МГУ им. М.В. Ломоносова
ЛЕКЦИЯ 1
ВВЕДЕНИЕ. МАТЕМАТИЧЕСКИЕ МОДЕЛИ В БИОЛОГИИ
Понятие модели. Объекты, цели и методы моделирования. Модели в разных науках. Компьютерные и математические модели. История первых моделей в биологии. Современная классификация моделей биологических процессов. Регрессионные, имитационные, качественные модели. Принципы имитацуионного моделирования и примеры моделей. Специфика моделирования живых систем.
Компьютеры в современном мире стали привычными для человеческой деятельности: в финансовой сфере, в бизнесе, промышленности, образовании, сфере досуга. Благодаря компьютерам западной цивилизации удалось существенно продвинуться в следующих направлениях.
- Автоматизация трудовой деятельности во всех сферах
- Информационная революция. Возможность хранить и структурировать огромные и самые разнообразные массивы информации и производить быстрый и эффективный поиск необходимой информации.
- Прогнозирование. Компьютер позволяет строить имитационные модели сложных систем, проигрывать сценарии и делать прогнозы.
- Оптимизация. Любая человеческая деятельность, в том числе обыденная жизнь требует постоянной оптимизации действий. В процессе эволюции сформировались биологические системы, которые оказываются оптимальными в том или ином смысле, например, в смысле наиболее экономичного использования энергии. Для того чтобы формализовать целевую функцию, то есть ответить на вопрос, что же является для системы оптимальным, необходимо сформулировать модель оптимизируемого процесса и критерии оптимизации. Компьютер позволяет проектировать и реализовать различные алгоритмы оптимизации.
Компьютер работает не с самой системой, а с моделью. Что же такое МОДЕЛЬ?
Наиболее простой и общий ответ на этот вопрос: модель — это копия объекта, в некотором смысле «более удобная», допускающая манипуляции в пространстве и во времени.
При моделировании, выборе и формулировке модели, определяющими обстоятельствами являются объект, цель и метод (средства) моделирования.
В нашем курсе объектами моделирования будут биологические процессы разного уровня организации.
Методами моделирования служат методы динамической теории систем. Средства — дифференциальные и разностные уравнения, методы качественной теории дифференциальных уравнений, компьютерная симуляция.
- Выяснение механизмов взаимодействия элементов системы
- Идентификация и верификация параметров модели по экспериментальным данным.
- Оценка устойчивости системы (модели). Само понятие устойчивости требует формализации.
- Прогноз поведения системы при различных внешних воздействиях, различных способах управления и проч.
- Оптимальное управление системой в соответствии с выбранным критерием оптимальности.
Примеры моделей.
1. Портрет дамы.
Пусть некто заказывает художнику написать портрет любимой женщины. Рассмотрим объект, метод (средства) и цель моделирования.
Объектом моделирования является женщина.
Метод (средства) — краски, кисти, холст. Эмаль, если портрет будет сделан на медальоне, как это было принято в прошлые века. Фотоаппарат и пленка. Рекламный щит, если некто хочет, чтобы его даму видели все, кто проезжает по оживленной магистрали. Обложка журнала, или экран телевизора. Наконец, сам художник, фотограф или рекламное агентство в лице своих дизайнеров.
Цель. При моделировании целью, как правило, является манипуляция с пространством и временем. Сохранить облик дамы во времени. Повесить портрет в гостиной, или медальон с изображением любимой — на шею, как это делали в старину. Чтобы потомки восхищались красотой дамы и своим пращуром, которому удалось запечатлеть такую красоту.
Другая цель — воспроизведение изображения (модели) объекта с целью сделать модель доступной некоторому кругу людей. Или многократно тиражировать, если некто хочет, чтобы образ дамы увидели миллионы.
2. Самолет в аэродинамической трубе. Помещая самолет в аэродинамическую трубу и испытывая его в различных воздушных потоках, мы решаем задачу взаимодействия системы с внешней средой. Это еще одна очень важная цель моделирования. При этом в корпусе самолета не обязательно должны находиться кресла, и тем более, стюардессы. Какие из свойств объекта необходимо учесть, а какие можно опустить, степень подробности воспроизведения моделью объекта, определяется теми вопросами, на которые хотят ответить с помощью модели.
3. Аквариум является примером физического моделирования. В аквариуме можно моделировать водную экосистему — речную, озерную, морскую, заселить ее некоторыми видами фито- и зоопланктона, рыбами, поддерживать определенный состав воды, температуру, даже течения. И строго контролировать условия эксперимента. Какие компоненты естественной системы будут воспроизведены, и с какой точностью, зависит от цели моделирования.
4. Выделенные из листьев хлоропласты. На выделенных системах часто изучают процессы, происходящие в живой системе, в этом смысле фрагмент является моделью целой живой системы. Выделение более простой системы позволяет исследовать механизмы процессов на молекулярном уровне. При этом исключается регуляция со стороны более высоких уровней организации, в данном случае, со стороны растительной клетки, листа, наконец, целого растения. В большинстве случаев наблюдать процессы на молекулярном уровне в нативной (ненарушенной) системе не представляется возможным. Говорят, что изученные на выделенном хлоропласте первичные процессы фотосинтеза являются моделью первичных процессов фотосинтеза в живом листе. К сожалению, этот метод фрагментирования приводит к тому, что «…живой ковер жизни распускается по ниточкам, каждая ниточка досконально изучается, но волшебный рисунок жизни оказывается утрачен» (лауреат Нобелевской премии по биохимии Л. Поллинг).
5. Бислойная липидная мембрана. Еще «более модельным» примером является изучение процессов ионного трансмембранного переноса на искусственной бислойной липидной мембране. Понятно, что в реальных биологических объектах мембраны чаще всего не бислойные, а многослойные, содержат встроенные белки и другие компоненты, поверхность их не является плоской и обладает множеством других индивидуальных особенностей. Однако, чтобы изучить законы образования поры, через которую ион проходит сквозь мембрану внутрь клетки или органеллы, необходимо создать «чистую», «модельную» систему, которую можно изучать экспериментально, и для которой можно использовать хорошо разработанное наукой физическое описание.
6. Популяция дрозофилы, является классическим объектом моделирования микроэволюционного процесса и примером исключительно удачно найденной модели. Еще более удобной моделью являются вирусы, которые можно размножать в пробирке. Хотя не вполне ясно, справедливы ли эволюционные закономерности, установленные на вирусах, для законов эволюции высших животных. В лекции 11 мы увидим, что хорошей моделью микроэволюционных процессов являются также микробные популяции в проточном культиваторе.
Из приведенных примеров видно, что любая физическая модель обладает конкретными свойствами физического объекта. В этом ее преимущества, но в этом и ее ограничения.
Компьютерные модели содержат «знания» об объекте в виде математических формул, таблиц, графиков, баз данных и знаний. Они позволяют изучать поведение системы при изменении внутренних характеристик и внешних условий, проигрывать сценарии, решать задачу оптимизации. Однако каждая компьютерная реализация соответствует конкретным, заданным параметрам системы. Наиболее общими и абстрактными являются математические модели.
Математические модели описывают целый класс процессов или явлений, которые обладают сходными свойствами, или являются изоморфными. Наука конца 20 века — синергетика, показала, что сходными уравнениями описываются процессы самоорганизации самой разной природы: от образования скоплений галактик до образования пятен планктона в океане.
Если удается сформулировать «хорошую» математическую модель, для ее исследования можно применить весь арсенал науки, накопленный за тысячелетия. Недаром многие классики независимо высказывали одну и ту же мудрую мысль:
«Область знания становится наукой, когда она выражает свои законы в виде математических соотношений»
С этой точки зрения самая «научная» наука ? физика. Она использует математику в качестве своего естественного языка. Все физические законы выражаются в виде математических формул или уравнений.
В химию математика пришла в тридцатые годы 20 века вместе с химической кинетикой и физической химией. Сейчас книги по химии, в особенности по химической кинетике, физической химии, квантовой химии полны математическими символами и уравнениями.
Чем более сложными являются объекты и процессы, которыми занимается наука, тем труднее найти математические абстракции, подходящие для описания этих объектов и процессов. В биологию, геологию и другие «описательные науки» математика пришла по настоящему только во второй половине 20 века.
Первые попытки математически описать биологические процессы относятся к моделям популяционной динамики. Эта область математической биологии и в дальнейшем служила математическим полигоном, на котором «отрабатывались» математические модели в разных областях биологии. В том числе модели эволюции, микробиологии, иммунологии и других областей, связанных с клеточными популяциями.
Самая первая известная модель, сформулированная в биологической постановке, ? знаменитый ряд Фибоначчи, который приводит в своем труде Леонардо из Пизы в 13 веке. Это ряд чисел, описывающий количество пар кроликов, которые рождаются каждый месяц, если кролики начинают размножаться со второго месяца и каждый месяц дают потомство в виде пары кроликов. Ряд представляет последовательность чисел:
Основные понятия моделей описываемых одним уравнением
МОДЕЛИ БИОЛОГИЧЕСКИХ СИСТЕМ, ОПИСЫВАЕМЫЕ
ОДНИМ ДИФФЕРЕНЦИАЛЬНЫМ УРАВНЕНИЕМ ПЕРВОГО ПОРЯДКА
Модели, приводящие к одному дифференциальному уравнению. Понятие решения одного автономного дифференциального уравнения. Стационарное состояние (состояние равновесия). Устойчивость состояния равновесия. Методы оценки устойчивости. Решение линейного дифференциального уравнения Примеры: экспоненциальный рост, логистический рост.
Изучение математических моделей биологических систем начнем с систем первого порядка, которым соответствует одно дифференциальное уравнение первого порядка:
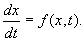
Если система автономная, то правая часть уравнений не зависит явно от времени и уравнение имеет вид:
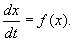 (2.1)
(2.1)
Состояние таких систем в каждый момент времени характеризуется одной единственной величиной – значением переменной x в данный момент времени t.
Рассмотрим плоскость t, x. Решениями уравнения (2.1): x( t) являются кривые на плоскости t, x , называемые интегральными кривыми (рис. 2.1)
Пусть заданы начальные условия 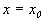 при t =0 или, иначе, пусть на плоскости t, x задана точка с координатами
при t =0 или, иначе, пусть на плоскости t, x задана точка с координатами 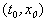 . Если для уравнения (2.1) выполнены условия теоремы Коши, то имеется единственное решение уравнения (2.1), удовлетворяющее этим начальным условиям, и через точку
. Если для уравнения (2.1) выполнены условия теоремы Коши, то имеется единственное решение уравнения (2.1), удовлетворяющее этим начальным условиям, и через точку 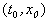 проходит одна единственная интегральная кривая x( t) .
проходит одна единственная интегральная кривая x( t) .
Рис. 2.1. Интегральные кривые x ( t ); – решения уравнения f ( x ) = 0

Интегральные кривые уравнения (2.1) не могут пересекаться. Решения уравнения (2.1) не могут быть периодическими, они монотонны.
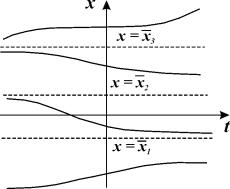
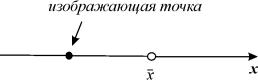
Поведение интегральных кривых на плоскости t, x можно установить, не решая в явном виде дифференциального уравнения (2.1), если известен характер движения изображающей точки на фазовой прямой.
Рассмотрим плоскость t, x , причем фазовую прямую совместим с осью x . Построим на плоскости t, x точку с абсциссой t и с ординатой, равной смещению изображающей точки по оси x в данный момент времени t. С течением времени в соответствии с уравнением (2.1) изображающая точка будет двигаться по фазовой прямой (рис. 2.2), а на плоскости t, x описывать некую кривую. Это будет интегральная кривая уравнения (2.1).
Решения одного автономного дифференциального уравнения либо уходят в бесконечность (чего не бывает в реальных системах), либо асимптотически приближаются к стационарному состоянию.
Стационарное состояние (точка покоя, особая точка, состояние равновесия)
В стационарном состоянии значения переменных в системе не меняются со временем. На языке дифференциальных уравнений это означает:
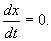 (2.2)
(2.2)
Если левая часть уравнения равна нулю, значит равна нулю и его правая часть:
Корни алгебраического уравнения (2.3): 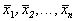 суть стационарные состояния дифференциального уравнения (2.1). На плоскости ( t, x) прямые
суть стационарные состояния дифференциального уравнения (2.1). На плоскости ( t, x) прямые 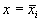 – асимптоты, к которым приближаются интегральные кривые. На фазовой прямой (рис. 2.2) стационарное состояние
– асимптоты, к которым приближаются интегральные кривые. На фазовой прямой (рис. 2.2) стационарное состояние 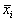 – точка, к которой стремится величина x.
– точка, к которой стремится величина x.
Реальные биологические системы испытывают многочисленные флуктуации, переменные при малых отклонениях возвращаются к своим стационарным значениям. Поэтому при построении модели важно знать, устойчивы ли стационарные состояния модели.
Рис. 2.3. К понятию устойчивости состояния равновесия

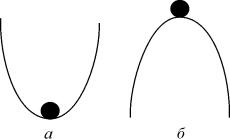
Устойчивость состояния равновесия
Каждый имеет интуитивное представление об устойчивости. На рис. 2.3. в обоих положениях (а и б) шарик находится в равновесии, т.к. сумма сил, действующих на него, равна нулю.
Попытайтесь ответить на вопрос : «Какое из этих состояний равновесия устойчиво?»
Скорее всего, Вы дали правильный ответ. Сказать, как Вы догадались? Вы дали шарику малое отклонение от состояния равновесия . В случае ( а) шарик вернулся. В случае ( б) покинул состояние равновесия навсегда.
Устойчивое состояние равновесия можно определить так: если при достаточно малом отклонении от положения равновесия система никогда не уйдет далеко от особой точки, то особая точка будет устойчивым состоянием равновесия, что соответствует устойчивому режиму функционирования системы.
Строгое математическое определение устойчивости состояния равновесия уравнения dx/dt = f( x) выглядит следующим образом :
Состояние равновесия устойчиво по Ляпунову, если задав сколь угодно малое положительное 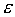 , всегда можно найти такое
, всегда можно найти такое 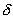 , что
, что
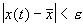 для
для 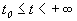 если
если 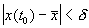 .
.
Иначе говоря, для устойчивого состояния равновесия справедливо утверждение: если в момент времени 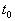 отклонение от состояния равновесия мало (
отклонение от состояния равновесия мало ( 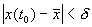 ), то в любой последующий момент времени
), то в любой последующий момент времени 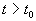 отклонение решения системы от состояния равновесия будет также мало:
отклонение решения системы от состояния равновесия будет также мало: 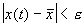 .
.
Другими словами: c тационарное состояние называется устойчивым, если малые отклонения не выводят систему слишком далеко из окрестности этого стационарного состояния. Пример — шарик в ямке (с трением или без трения).
Стационарное состояние называется асимптотически устойчивым, если малые отклонения от него со временем затухают. Пример — шарик в ямке в вязкой среде.
Стационарное состояние называется неустойчивым, если малые отклонения со временем увеличиваются. Пример: шарик на горке.
Устойчивое стационарное состояние представляет собой простейший тип аттрактора.
Аттрактором называется множество, к которому стремится изображающая точка системы с течением времени (притягивающее множество).
В нашем курсе мы рассмотрим следующие типы аттракторов:
· устойчивая точка покоя;
· предельный цикл — режим колебаний с постоянными периодом и амплитудой (начиная с размерности системы 2 );
· Области с квазистохастическим поведением траекторий в области аттрактора, например, «странный аттрактор» (начиная с размерности 3 ).
Аналитический метод исследования устойчивости стационарного состояния (метод Ляпунова). Линеаризация системы в окрестности стационарного состояния.
Метод Ляпунова приложим к широкому классу систем различной размерности, точечным системам, которые описываются обыкновенными дифференциальными уравнениями, и распределенным системам, описываемым уравнениями в частных производных, непрерывным и дискретным.
Рассмотрим метод линеаризации Ляпунова для одного автономного дифференциального уравнения первого порядка. Пусть 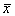 — стационарное решение уравнения:
— стационарное решение уравнения:
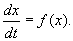 (2.1)
(2.1)
Пусть система, первоначально находившаяся в стационарном состоянии, отклонилась от него и перешла в близкую точку с координатой: 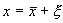 , причем
, причем 
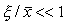 .
.
Перейдем в уравнении (2.1) от переменной x к переменной 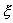 , т.е. новой переменной будет отклонение системы от стационарного состояния.
, т.е. новой переменной будет отклонение системы от стационарного состояния.
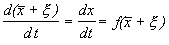 .
.
Учтем, что 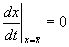 по определению стационарного состояния.
по определению стационарного состояния.
Правую часть разложим в ряд Тейлора в точке 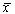 :
:
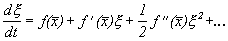
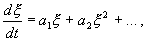
где 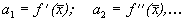
Отбросим члены порядка 2 и выше. Останется линейное уравнение:
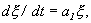 (2.4)
(2.4)
которое носит название линеаризованного уравнения или уравнения первого приближения. Интеграл этого уравнения для 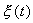 находится сразу:
находится сразу:
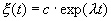 , (2.5)
, (2.5)
где 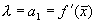 , с — произвольная постоянная.
, с — произвольная постоянная.
Если SYMBOL 108 \f «Symbol» \s 12 l SYMBOL 60 \f «Symbol» \s 12 0 , то при 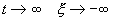 и, следовательно, первоначальное отклонение SYMBOL 120 \f «Symbol» \s 12 x от состояния равновесия со временем затухает. Это означает, по определению, что состояние равновесия устойчиво.
и, следовательно, первоначальное отклонение SYMBOL 120 \f «Symbol» \s 12 x от состояния равновесия со временем затухает. Это означает, по определению, что состояние равновесия устойчиво.
Если же SYMBOL 108 \f «Symbol» \s 12 l SYMBOL 62 \f «Symbol» \s 12 > 0 , то при 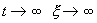 , и исходное состояние равновесия неустойчиво.
, и исходное состояние равновесия неустойчиво.
Если SYMBOL 108 \f «Symbol» \s 12 l =0 , то уравнение первого приближения не может дать ответа на вопрос об устойчивости состояния равновесия системы. Необходимо рассматривать члены более высокого порядка в разложении в ряд Тейлора. Такие случаи мы рассмотрим в лекции 6.
Аналогичные рассуждения проводятся при рассмотрении устойчивости стационарных состояний более сложных динамических систем.
Итак, устойчивость стационарного состояния 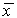 уравнения dx/dt=f(x) определяется знаком производной правой части в стационарной точке.
уравнения dx/dt=f(x) определяется знаком производной правой части в стационарной точке.
В случае одного уравнения вопрос об устойчивости состояния равновесия нетрудно решить, рассматривая график функции f(x).
По определению в стационарной точке правая часть уравнения (2.1) ‑ функция f(x) обращается в нуль.
Здесь возможны три случая (рис. 2.4 а, б, в).
1. Вблизи состояния равновесия функция f(x) меняет знак с плюса на минус при возрастании x (рис. 2.4 а).
Отклоним изображающую точку системы в сторону 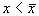 . В этой области скорость изменения x dx/dt = f(x) положительна. Следовательно, x увеличивается, т.е. возвращается к
. В этой области скорость изменения x dx/dt = f(x) положительна. Следовательно, x увеличивается, т.е. возвращается к 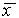 . При
. При 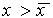 скорость изменения величины x уменьшается, т.к. функция f(x) SYMBOL 60 \f «Symbol» \s 12 0. Следовательно, здесь x уменьшается и опять стремится к
скорость изменения величины x уменьшается, т.к. функция f(x) SYMBOL 60 \f «Symbol» \s 12 0. Следовательно, здесь x уменьшается и опять стремится к 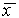 . Таким образом, отклонения от стационарного состояния в обе стороны затухают. Стационарное состояние устойчиво.
. Таким образом, отклонения от стационарного состояния в обе стороны затухают. Стационарное состояние устойчиво.
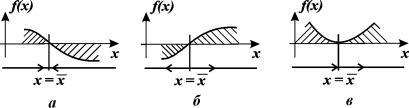
Рис. 2.4. Определение устойчивости стационарного состояния по графику функции f( x)
a – стационарное состояние 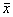 устойчиво;
устойчиво;
б, в ‑ стационарное состояние 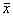 неустойчиво
неустойчиво
2. Вблизи состояния равновесия функция f ( x) меняет знак с минуса на плюс при возрастании x ( рис. 2.4 б) .
Проведите рассуждения, аналогичные случаю 1. Поместите изображающую точку в область 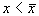 . Теперь в область
. Теперь в область 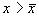 .
.
В обоих случаях изображающая точка удаляется от состояния равновесия. Стационарное состояние неустойчиво.
3. Вблизи состояния равновесия функции f(x) не меняет знак ( рис 2.4 в) .
Поскольку 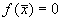 , это означает, что изображающая точка, помещенная достаточно близко к состоянию равновесия с одной стороны, будет приближаться к нему, помещенная с другой стороны – удаляться.
, это означает, что изображающая точка, помещенная достаточно близко к состоянию равновесия с одной стороны, будет приближаться к нему, помещенная с другой стороны – удаляться.
Вопрос. Является ли состояние равновесия в случае 3 устойчивым?
Ответ. Нет. По определению устойчивости.
1. Рост колонии микроорганизмов
За время D t прирост численности равен:
где R – число родившихся и S – число умерших за время SYMBOL 68 \f «Symbol» \s 12 D t особей пропорциональные этому промежутку времени:

В дискретной форме:
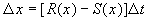 .
.
Разделив на SYMBOL 68 \f «Symbol» \s 12 D t и переходя к пределу при t SYMBOL 174 \f «Symbol» \s 12 ® 0 , получим дифференциальное уравнение
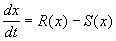 . (2.6)
. (2.6)
В простейшем случае, когда рождаемость и смертность пропорциональны численности:
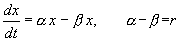 ,
,
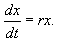 (2.7)
(2.7)
Разделим переменные и проинтегрируем:
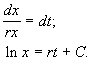
Переходя от логарифмов к значениям переменной x и определяя произвольную постоянную С из начальных условий, получим экспоненциальную форму динамики роста.
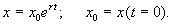 (2.8)
(2.8)
График функции (2.8) при положительных (размножение) и отрицательных (вымирание) значениях константы скорости роста представлен на рис. 2.5. Роль этой модели в развитии математической биологии и экологии мы обсудим в Лекции 3.
Рис. 2.5. Экспоненциальная форма динамики роста численности колонии микроорганизмов в соответствии с системой уравнений (2.7)

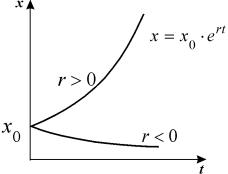
2. Вещество переходит в раствор
Пусть количество вещества, переходящего в раствор, пропорционально интервалу времени и разности между максимально возможной концентрацией Р и концентрацией x в данный момент времени: 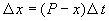 .
.
В форме дифференциального уравнения этот закон выглядит в
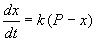 . (2.9)
. (2.9)
Разделим в этом уравнении переменные, и проинтегрируем:
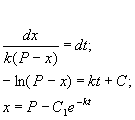 (2.10)
(2.10)
Здесь C 1 — произвольная постоянная. Если x (0) = 0,
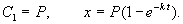
График этой функции представлен на рис. 2.6. – он представляет собой кривую с насыщением.
Рис. 2.6. Концентрация вещества х в зависимости от времени. График уравнения 2.9.

Какие дифференциальные уравнения можно решать аналитически?
Лишь для ограниченных классов дифференциальных уравнений разработаны аналитические методы решения. Подробно они изучаются в курсах дифференциальных уравнений. Отметим основные из них/
1. Уравнения с разделяющимися переменными решаются в интегралах. К ним относятся оба приведенные выше примера.
2. Линейные дифференциальные уравнения (не обязательно автономные).
3. Некоторые специальные виды уравнений.
Решение линейного уравнения
Линейным дифференциальным уравнением 1-го порядка называют уравнение, линейное относительно искомой функции и ее производной. Оно имеет вид:
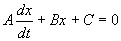 . (2.11)
. (2.11)
Здесь A, B, C — заданные непрерывные функции от t.
Пусть в некотором интервале изменения t A SYMBOL 185 \f «Symbol» \s 12 _ 0 . Тогда на него можно разделить все члены уравнения. При этом получим:
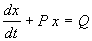 . (2.12)
. (2.12)
Eсли Q=0 , уравнение (2.12) называется однородным, если Q SYMBOL 185 \f «Symbol» \s 12 _ 0 – неоднородным.
Решим сначала однородное уравнение.
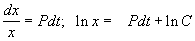 .
.
Общее решение линейного однородного уравнения имеет вид:
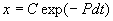 . (2.13)
. (2.13)
Чтобы найти решение неоднородного уравнения применим метод вариации постоянной. Будем считать С неизвестной функцией t . Подставляя правую часть выражения (2.13) в уравнение (2.12), имеем:
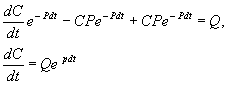
Теперь С находим интегрированием: 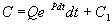 . Здесь С1 – произвольная постоянная.
. Здесь С1 – произвольная постоянная.
Итак, общее решение линейного неоднородного уравнения первого порядка:
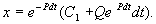 (2.14)
(2.14)
Таким образом, решение уравнения (2.12) представляет собой сумму двух слагаемых:
1) общее решение однородного уравнения (2.13) и
2) частное решение неоднородного уравнения, которое получается из общего решения, если С1 = 0.
Рассмотрим еще один пример, который относится к классическим моделям математической экологии. Логистическое уравнение было предложено Ферхюльстом в 1838 г. Оно имеет вид:
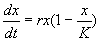 . (2.15)
. (2.15)
Это уравнение обладает двумя важными свойствами. При малых х численность х возрастает, при больших – приближается к определенному пределу К .
Уравнение (2.15) можно решить аналитически. Ход решения следующий. Произведем разделение переменных:
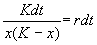 . (2.16)
. (2.16)
Представим левую часть в виде суммы и проинтегрируем
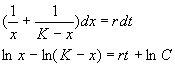
Переходя от логарифмов к переменным, получим:
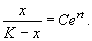 (2.17)
(2.17)
Здесь С – произвольная постоянная, которая определяется начальным значением численности x0 :
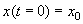 ;
; 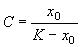 .
.
Подставим это значение С в формулу (2.17):
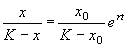 .
.
Отсюда получим решение – зависимость численности от времени:
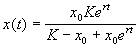 . (2.18)
. (2.18)
График функции (2.18) при разных начальных значениях численности популяции представлен на рис. 2.7.
Рис.2.7. Динамика численности в логистической модели 2.18
при разных начальных значениях численности

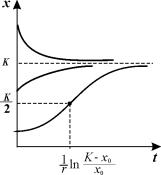
Если начальное значение х0 К/2, кривая роста имеет точку перегиба. Если х0 > К, численность со временем убывает.
В приведенных примерах в правой части уравнений стоят полиномы первой и второй степени. Если в правой части ‑ более сложная нелинейная функция, алгебраическое уравнение для стационарных значений может иметь несколько корней. Какое из этих решений реализуется в этом случае, будет зависеть от начальных условий.
В дальнейшем мы, как правило, не будем искать аналитическое решение для наших моделей. Для более сложных нелинейных уравнений это и невозможно. Однако важные заключения относительно свойств моделей можно сделать и на основании качественного их исследования, в первую очередь путем исследования устойчивости стационарных состояний и типов поведения системы вблизи этих состояний. При этом следует иметь в виду, что с помощью одного автономного дифференциального уравнения могут быть описаны только монотонные изменения переменной, и, следовательно, ни периодические, ни хаотические процессы не могут быть описаны. Для описания более сложного поведения необходимо либо переходить к системам большей размерности (2, 3 порядка и выше), либо вводить время в явном виде в правую часть уравнения. В Лекции 3 мы увидим, что дискретные уравнения и уравнения с запаздыванием могут описать и колебания, и динамический хаос.
http://spkurdyumov.ru/education/kurs-lekcij-matematicheskie-modeli-v-biologii/
http://www.library.biophys.msu.ru/LectMB/Lect02.htm