Параграфы 32 и 34 Гликолиз. Аэробный обмен углевод
Автор текста – Анисимова Е.С.
Продавать текст нельзя. Авторские права защищены.
Для замечаний:
https://vk.com/bch_5
Параграфы 32 и 34.
Реакции, формулы и ферменты см. в файле «32 ФОРМУЛЫ ГЛИКОЛИЗа».
ПАРАГРАФ 32:
«Гликолиз».
Параграф 34:
«Аэробный обмен углеводов».
Гликолиз – это процесс превращения глюкозы в пируват (10 реакций).
Считается вторым этапом катаболизма (второй называется унификацией) в обмене углеводов – см. п.20.
Локализация процесса – гликолиз протекает во всех клетках организма, в гиалоплазме.
Глюкоза для гликолиза и других процессов поступает в клетки из крови, для чего в крови должна поддерживаться достаточная концентрация глюкозы (от 3 ммоль/л – см. п.37).
Транспорт глюкозы через мембраны клеток осуществляется специальными белками-переносчиками глюкозы. Транспорт глюкозы в клетки многих тканей (например, жировой и мышечной тканей, но не нейронов и эритроцитов) стимулируется инсулином (см. п.102, 103); и поэтому при снижении количества инсулина в крови или снижении действия инсулина (при диабете) транспорт глюкозы в клетки названных тканей снижен, что приводит к дефициту глюкозы в клетках и к избытку глюкозы в крови – см. п.37.
Кроме того, при расщеплении гликогена в мышцах (гликогенолизе – см.п.31) образуется метаболит гликолиза – глюкозо-6-фосфат, который сразу вступает во вторую реакцию гликолиза.
32.2. Конечные метаболиты гликолиза, значение и тканевые особенности
(то есть в каких клетках и тканях какие варианты бывают).
Центральным конечным метаболитом гликолиза является ПИРУВАТ (2 молекулы пирувата образуются из одной молекулы глюкозы).
Дальнейшие превращения пирувата зависят от условий, в которых протекает гликолиз – или в лактат, или в ацетилКоА, или в оксалоацетат (см. далее).
32.2.1. Превращения пирувата В АЭРОБНЫХ УСЛОВИЯХ
(АЭРОБНЫЙ ОБМЕН УГЛЕВОДОВ:
это гликолиз до пирувата и дальнейшие превращения пирувата, о которых тут сказано):
При наличии кислорода и возможности его использования (то есть в аэробных условиях в митохондриях во всех клетках, кроме эритроцитов)
1 – пируват может превратиться в АЦЕТИЛ-КоА под действием ПДГ (см. п.20, 21, «32 формулы гликолиз»), что даёт по 2,5 АТФ от каждой молекулы пирувата.
Пируват ; ацетилКоА + 2,5 АТФ
2.1 – АцетилКоА может поступить в ЦТК, что может привести к выработке 10 молекул АТФ от каждой молекулы ацетилКоА. И превращению ацетилКоА в углекислый газ для выдоха. Что завершает распад углеводородного скелета молекулы глюкозы.
ацетилКоА ; 2СО2 + 10 АТФ (при поступлении в ЦТК и ДЦ)
2.2 – В печени ацетилКоА может использоваться для синтеза жирных кислот (при сытости, см. п.46),кетоновых тел (при голоде и стрессе, см. п.47) и холестерина (см. п.49), а также (через реакции ЦТК) – аминокислот глутамата, глутамина и т.д. (см. п.67).
ацетилКоА ; жирные кислоты, кетоновые тела, холестерин, аминокислоты (с оксалоацетатом)
3 – Из пирувата может образоваться аминокислота АЛАНИН (см. п.64, 67 и 33) и кетокислота ОКСАЛОАЦЕТАТ (см. п.33), которая может 1) использоваться в ЦТК (см.п. 21) и 2) для синтеза аминокислоты аспартата для синтеза аспарагина, белков, для обезвреживания аммиака в синтезе мочевины (см.п.66) и для синтеза нуклеотидов (а затем РНК и ДНК).
4 – из метаболита гликолиза 3-фосфоглицерата (см. п.67) может синтезироваться аминокислота СЕРИН, из которой могут синтезироваться глицин, цистеин, липоиды (см. п.52).
Пируват ; аланин или оксалоацетат (; аспартат ; мочевина или нуклеотиды ; ДНК и РНК)
ОКСАЛОАЦЕТАТ для ЦТК или ; аспартат ; мочевина или нуклеотиды ; ДНК и РНК)
; (биотин!)
Аланин ; ПИРУВАТ ; (ПДГ) АЦЕТИЛ-КоА (+ 2,5 АТФ) ; (в ЦТК) 2СО2 + 10 АТФ (всего 12,5 АТФ)
; (под действием ПДГ, витамины – В1, В2, РР, пантотенат, липоевая)
ацетилКоА ; жирные кислоты (для жира и липоидов), кетоновые тела, холестерин
ЗНАЧЕНИЕ ГЛИКОЛИЗА в аэробных условиях – в получении всех перечисленных веществ, в том числе для синтезов (можно сказать, что это анаболическая роль гликолиза по аналогии с анаэробной ролью ЦТК). Участие гликолиза в выработке АТФ – это энергетическая роль гликолиза. Участие гликолиза в катаболизме глюкозы (превращение 6-тиуглеродной молекулы глюкозы в две трёхуглеродных молекулы пирувата) и поставка метаболитов для ЦТК (ацетилКоА и оксалоацетата) – катаболическая. Это достоинства гликолиза в аэробных условиях.
Недостатком аэробного обмена глюкозы является то, что его реакции протекают только в аэробных условиях и не протекают в анаэробных.
32.2.2. Превращения пирувата В АНАЭРОБНЫХ УСЛОВИЯХ.
При отсутствии кислорода или невозможности его использования в дыхательной цепи (см. п.22), то есть в анаэробных условиях пируват превращается в ЛАКТАТ (под действием фермента ЛДГ),
поскольку в анаэробных условиях пируват не может превратиться в ацетилКоА, так как не работает ПДГ (из-за избытка НАДН и дефицита НАД).
Причиной сниженной активности ПДГ и образования лактата может быть и дефицит витаминов ПДГ, особенно В1.
Лактат – это анион молочной кислоты, которая при клеточном рН (около 7) диссоциирует на лактат и протон; повышение концентрации протонов (Н+) называется ацидозом («закислением»), поскольку именно наличие протонов создаёт кислую среду, кислую реакцию, снижает рН.
Использовать кислород в дыхательной цепи не могут ЭРИТРОЦИТЫ из-за отсутствия в них митохондрий и дыхательной цепи,
а также любые клетки, если в них не работает дыхательная цепь (см.п.22) из-за попадания в них ингибиторов дыхательной цепи.
Дефицит кислорода (гипоксия) возникает во всех клетках при нарушении доставки кислорода:
1) из-за нарушения кровотока (из-за тромбов, жгута, сдавливания, падения артериального давления, большой кровопотери, сердечной недостаточности и т.д.),
2) из-за нарушения работы эритроцитов (см. п.121),
3) из-за нарушения поступления кислорода в лёгкие (при эмфиземе лёгких у курильщиков, при бронхоспазме и т.д.).
Превращение глюкозы в лактат в анаэробных условиях могут называть анаэробным гликолизом.
Лактат опасен, поскольку его накопление приводит к АЦИДОЗУ, который называется лактацидозом (в отличие от кетоацидоза при накоплении кетоновых тел – см. п. 47). Лактацидоз и кетоацидоз относятся к метаболическим ацидозам, то есть к ацидозам, обусловленным накоплением метаболитов (кислот) и нарушением метаболизма. Есть ещё респираторный ацидоз – то есть связанный с нарушением дыхания (задержкой дыхания). При всех ацидозах портится самочувствие, нарушается работа белков (так как оптимум для большинства белков – рН около 7), возникает чувство тошноты. Сильный ацидоз может привести к коме, но для здоровых людей это не характерно.
32.3. ОБМЕН ЛАКТАТА.
Это процессы образования и использования лактата.
32.3.1. ОБРАЗОВАНИЕ лактата.
Как сказано выше, образуется лактат при анаэробном гликолизе в эритроцитах, мышцах и других клетках в анаэробных условиях. Из клеток он может поступать в кровь, что при избытке лактата приводит к ацидозу.
Ткани-клетки-источники лактата:
В покое основное количество лактата поступает в кровь из эритроцитов, а при наряжённой работе мышц – из мышц.
Кроме этого, лактат поступает в кровь из любых клеток, которым не хватает кислорода или в которых не работают дыхательная цепь, ЦТК и ПДГ. Причиной сниженной работы ЦТК бывает дефицит витаминов ЦТК, особенно В1. См. 32.2.2.
32.3.2. Использование и УТИЛИЗАЦИЯ ЛАКТАТА.
Из крови большая часть лактата поступает в клетки печени, где утилизируется. Это снижает ацидоз. Поэтому нарушение работы печени (печёночная недостаточность) может стать причиной избытка лактата в крови (из-за сниженного использования лактата клетками печени).
Пути метаболизма лактата – 25% и 75%.
25% лактата подвергаются так называемому ОКИСЛЕНИЮ ЛАКТАТА.
При этом:
1) лактат превращается в пируват под действием фермента ЛДГ, что даёт 2,5 АТФ благодаря образованию НАДН и его использованию в дыхательной цепи,
2) затем пируват превращается в ацетилКоА под действием ПДГ, что даёт ещё 2,5 АТФ благодаря образованию НАДН и его использованию в дыхательной цепи,
3) затем ацетилКоА вступает в ЦТК, что даёт ещё 10 АТФ.
В сумме окисление одной молекулы лактата даёт 15 молекул АТФ.
Реакции окисления лактата идут только в аэробных условиях.
75% лактата используются для синтеза глюкозы, который называется глюконеогенезом (ГНГ) и протекает в основном в печени.
Поэтому снижение ГНГ может стать причиной избытка лактата и ацидоза.
При превращении лактата в глюкозу:
1) сначала лактат превращается в пируват под действием ЛДГ, как и при окислении лактата,
2) затем пируват превращается в ОКСАЛОАЦЕТАТ (см. п.33 и п.21), что требует витамина БИОТИН – без биотина реакция не идёт (причиной дефицита биотина, нарушения этой реакции, ГНГ и избытка лактата бывает избыточное употребление сырых яиц, так как в них содержится вещество, препятствующее усвоению биотина),
3) оксалоацетат превращается в глюкозу в ходе реакций глюконеогенеза.
32.3.3. Избыток лактата.
Между процессами образования лактата и его использования должно быть равновесие, благодаря которому концентрация лактата в клетках и крови не превышала бы нормальную.
Повышение концентрации лактата в клетках приводит к снижению рН в клетках, нарушению работы белков клеток, повреждению клеток. Например, мышцы при избыточной концентрации лактата теряют способность сокращаться.
Повышение концентрации лактата в крови называется гиперЛАКТАТемией и приводит к ацидозу.
К избытку лактата приводит или повышение образования лактата, или снижение утилизация лактата.
Причины повышенного образования лактата (см. выше 32.2.3. и 32.3) –
1) дефицит кислорода (гипоксия при анемии, сердечной или лёгочной недостаточности и т.д.) или снижение его использования,
2) снижение превращения пирувата в ацетилКоА из-за нарушения работы ПДГ при дефиците В1 или других витаминов ПДГ,
3) повышенная концентрация НАДН, которая бывает не только при дефиците кислорода, но и при употреблении алкоголя.
Причины сниженного использования лактата и пирувата:
1) дефицит витаминов ПДГ и биотина, без которых не протекают реакции использования и пирувата,
2) нарушение работы печени, в котором должен протекать ГНГ.
Для коррекции ацидоза, обусловленного избытком лактата, нужно убрать причины избытка лактата – восстановить кровоток, принимать витамины, не употреблять алкоголь и т.д.
32.4. Выработка АТФ при анаэробном гликолизе
(энергетический баланс анаэробного гликолиза).
В анаэробных условиях гликолиз (при котором глюкоза превращается в лактат) позволяет синтезировать 2 молекулы АТФ способом субстратного фосфорилирования (см. файл «32 формулы гликолиз» реакции № 7 и 10, а также п.23) в расчёте на использование одной молекулы глюкозы (превращение одной глюкозы в 2 молекулы лактата).
1. Этого количества хватает только эритроцитам (у них и нет других источников АТФ).
Остальным клеткам этого (2 молекул АТФ от одной молекулы глюкозы) мало, но без анаэробного гликолиза клетки не имели бы и этого в анаэробных условиях.
2. При наложении жгута и в других ситуациях нарушения кровотока ткани конечностей не подвергаются в течение часа некрозу именно благодаря тому, что есть небольшое количество АТФ, которое вырабатывается при гликолизе в анаэробных условиях. А после часа времени возникает риск некроза именно потому, что АТФ от анаэробного гликолиза мало.
3. Клеткам головного мозга недостаточно того количества АТФ, который может им дать анаэробный гликолиз, поэтому они не способны, в отличие от тканей конечностей, выдержать гипоксию в течение часа.
4. При интенсивной работе мышц (например, при беге на сто метров или при подъёме штанги) в них возникают анаэробные условия. Наличие в них АТФ для работы поддерживается именно благодаря 2 АТФ анаэробного гликолиза, который быстро «включается» при работе мышц.
Ранее анаэробного гликолиза АТФ образуется благодаря запасам креатин/фосфата – см. п.68. Более длительная работа мышц требует получения АТФ при аэробных процессах – аэробном обмене глюкозы и при ;-окислении жирных кислот.
32.5. Достоинства и недостатки анаэробного гликолиза.
Таким образом, то, что анаэробный гликолиз даёт хоть немного АТФ без кислорода – это его ДОСТОИНСТВО.
А то, что этого количества АТФ МАЛО (хватает только эритроцитам, тканям конечностей при гипоксии и скелетным мышцам при короткой напряжённой работе, но не хватает другим тканям вообще, а тканям конечностей не хватает более чем на час, не хватает мышцам для длительной работы – это НЕДОСТАТОК анаэробного гликолиза).
Ещё один недостаток анаэробного гликолиза – то, что избыток его конечного метаболита (лактата) приводит к ацидозу.
Гликолиз является единственным процессом, за счёт которого может образоваться АТФ при отсутствии (или неиспользовании) кислорода. Именно гликолиз обеспечивает молекулами АТФ эритроциты, в которых нет других источников АТФ из-за отсутствия митохондрий и дыхательной цепи см. п.23.
32.6. Выработка АТФ при аэробном обмене глюкозы.
См. «32 формулы гликолиз» и 32.2.
В аэробных условиях глюкоза превращается в 6 молекул СО2, что сопровождается синтезом 32 молекул АТФ.
При гликолизе глюкоза превращается в 2 молекулы пирувата, что сопровождается образованием 2 молекул АТФ способом субстратного фосфорилирования и двух молекул НАДН (см. реакцию гликолиза № 6).
Каждая из этих 2 молекул НАДН даёт по 2,5 АТФ, то есть всего 5 (с помощью ДЦ).
Каждая из двух молекул пирувата даёт по 12,5 молекул АТФ, то есть всего – 25.
В сумме глюкоза может дать 32 молекулы АТФ. (2 + 2,5*2 + 12,5*2).
В головном мозге из-за особенностей обмена глюкоза даёт не 32, а 30 молекул АТФ.
32.7. РЕГУЛЯЦИЯ ГЛИКОЛИЗА. См. п.6 и 7, 33, 37, 102.
Смысл регуляции любого процесса в том, чтобы его скорость соответствовала потребностям организма и отдельных клеток.
Изменение скорости процесса осуществляется через изменение активности или концентрации ферментов процесса под влиянием метаболитов и гормонов.
Активность повышается активаторами и снижается ингибиторами.
Концентрация ферментов повышается в результате усиления синтеза ферментов под влиянием индукторов и снижается в результате снижения синтеза под влиянием репрессоров.
Регулировать все ферменты 10 реакций гликолиза не обязательно. Достаточно регулировать активность некоторых ферментов. Эти регулируемые ферменты называются КЛЮЧЕВЫМИ – см. п.7.
В гликолизе ключевыми ферментами являются три – ферменты первой реакции, последней и третьей – гексокиназа, фосфофруктокиназа и пируваткиназа.
Фермент первой реакции последовательности реакций часто является ключевым. Фермент третьей реакции гликолиза является самым медленным – это тоже типично (что фермент самой медленной реакции метаболического пути является ключевым).
32.7.1. Регуляция гликолиза гормонами.
Гликолиз стимулируется инсулином (за счёт индукции ключевых ферментов) и подавляется глюкокортикостероидами (ГКС; за счёт репрессии ключевых ферментов). См. п.85.
Катехоламины адреналин и норадреналин усиливают гликолиз в мышцах и подавляют в печени.
Благодаря стимуляции инсулином гликолиза (а также стимуляции инсулином транспорта глюкозы во многие клетки) расход глюкозы тканями увеличивается, что приводит
1) к удовлетворению потребностей клеток в глюкозе и
2) к снижению концентрации глюкозы в крови –
это снижение концентрации глюкозы в крови называется гипогликемическим эффектом,
а инсулин, который обусловливает снижение концентрации глюкозы в крови, является гипогликемическим гормоном.
32.7.2. Регуляция гликолиза метаболитами.
Обычно активаторами процесса являются его субстраты, а ингибиторами – продуктами. Механизм – аллостерический (см. п.6).
Активатором гликолиза является АДФ (в качестве субстрата – чтобы превратиться в АТФ), а ингибиторами – продукты гликолиза АТФ, ацетилКоА и цитрат (отдалённо это продукты гликолиза, так как цитрат образуется из ацетилКоА и оксалоацетата, образующихся из пирувата).
Эффекты АДФ противоположны эффектам АТФ, так как АДФ образуется при расщеплении АТФ, и [АДФ] становится тем больше, чем меньше [АТФ].
Ещё один активатор гликолиза – метаболит фруктозо-2,6- бисфосфат, который активирует фосфофруктокиназу.
Сравните регуляцию гликолиза с регуляцией ГНГ – эти два процесса регулируются одними и теми же регуляторами, но действие регуляторов на эти два противоположных процесса прямо противоположно. Это помогает рассказать регуляцию ГНГ, зная регуляцию гликолиза.
Р е г у л я ц и я г о р м о н а м и процессов углеводного обмена.
Усиление процесса
способствует (гипер/гликемии) или не способствует? Г о р м о н ы
Ин ГГ КА ГКС
; [Г] Гипергликемические
Гликолиз (глюкоза в пируват) Нет, ведёт к гипогликемии ; ; в печени см. *
Глюконеогенез (синтез глюкозы) Да, ведёт к гипергликемии ; ; в печени
* В мышцах КА (в т.ч. адреналин) не снижают гликолиз, а усиливают – так как КА должны обеспечить мышцы энергией за счет выработки в мышцах АТФ. Гликолиз – это единственный процесс, который может дать АТФ при дефиците кислорода (дефицит кислорода обязательно возникает в работающих скелетных мышцах, как бы ни старались легкие и сердце обеспечить мышцы кислородом; но чем лучше человек тренирован, тем лучше организм справляется с нагрузками).
Р е г у л я ц и я гликолиза и ГНГ метаболитами. (Аллостерическая).
(Действие регуляторов на гликолиз и на ГНГ противоположно.)
ацетилКоА Цитрат А Т Ф А Д Ф Фруктозо-2,6-бисфосфат
гликолиз Продукт продукт Продукт Субстрат
Ингибиторы гликолиза Активаторы гликолиза
ГНГ: синтез
глюкозы Субстрат продукт
Активаторы ГНГ Ингибиторы ГНГ
Таблица «Гликолиз в разных условиях: в анаэробных и в аэробных». ;
Гликолиз – это «лизис глюкозы», то есть расщепление (катаболизм) глюкозы (ее углеродного скелета) до пирувата,
а далее – в зависимости от условий: в анаэробных условиях пируват превращается в лактат,
а в аэробных поступает в ЦТК и превращается в СО2 и воду (с участием ДЦ).
+ — достоинство процесса, – недостаток.
№ В анаэробных В аэробных
1. Требует ли н а л и ч и я кислорода Нет (это +) Да (это – )
2. Может ли протекать при ишемии, гипоксии (1) Да (это +) нет
3. Нужна ли помощь ЦТК и ДЦ нет да
4. Каким метаболитом заканчивается Лактат (снижает рН, это –) 6 СО2, НОН
5. Приводит ли к образованию лактата Да (это –) Нет (это +)
6. Приводит ли к ацидозу при усилении (см. 5) Да (это –) Нет (это +)
7. Нужно ли у ч а с т и е митохондрий Нет Да
8. Протекает ли в эритроцитах (см. 7) да Нет (т.к. в ЭЦ нет МХ)
9. Сколько АТФ дает (в расчете на 1 молекулу Г) 2 АТФ 32 АТФ (30 в мозге)
10 Достаточен ли для мозга
в качестве источника энергии Нет (поэтому мозг
без О2 гибнет) Да (основной источник
АТФ для мозга)
11 Дает ли метаболиты для ЦТК (ОА и др.) Нет (это –) да (это +)
12
В чем д о с т о и н с т в а
(см. выше) 1) только он дает АТФ в ЭЦ,
2) только он дает АТФ без О2, хоть и мало (всего 2),
3) быстро включается, что ценно при короткой напряженной работе 1) дает много АТФ,
2) дает метаболиты ЦТК
(ОА и ацетилКоА из П,
а затем в ЦТК образуются остальные, в т.ч. для аминокислот и белков)
В чем н е д о с т а т к и
(см. выше) 1) образует лактат, что приводит к ацидозу,
2) мало АТФ (из-за чего жгут нельзя долго держать) не протекает
без кислорода
(при гипоксии)
14 Каким
тканям или клеткам
дает достаточное количество энергии Эритроцитам, скелетным мышцам при короткой работе, а при жгуте – час Всем, в которых протекает
(в эритроцитах не протекает)
О б м е н л а к т а т а . Лактат (Л) – это соль или анион молочной кислоты.
1. Как называется увеличение [Л] в крови Гипер / лактат / емия
2. Чем о п а с н а высокая [ лактата ]
(молочная кислота ; лактат + Н+ ) Накоплением протонов (Н+), т.е. АЦИДОЗОМ, который приводит к
тошноте, усталости и (при очень сильном ацидозе) к коме
3. Какой ПРОЦЕСС образует лактат Анаэробный гликолиз (при дефиците кислорода и в ЭЦ)
4. Какие ТКАНИ являются источником лактата (поступающего в кровь) В покое – эритроциты. При работе скелетных мышц – мышцы.
При патологии – любая ткань при гипоксии и без В1.
5. Какие ПРОЦЕССЫ используют лактат ГНГ (в печени) и окисление лактата (ЛДГ, ПДГ, ЦТК)
6. Какой ОРГАН «берёт» лактат из крови Печень (для превращения лактата в глюкозу в ГНГ)
7.
Причины накопления лактата
(гипер/лактат/емии),
связанные
с увеличением образования лактата
(см. 3 и 4) У здоровых – работа скелетных мышц (усталость, тошнота).
У больных – разные формы Г И П О К С И И , в т.ч:
1. из-за ишемии (жгут, тромб, спазм, падение АД и т.д.),
2. из-за патологии дыхательной системы (эмфизема легких при курении и др.),
3. из-за патологии сердечно-сосудистой системы,
4. из-за анемии (нет Fe, витаминов, белка, есть гемолиз),
5. при шоке, коллапсе, терминальных состояниях.
8. Причины накопления лактата,
связанные с уменьшением его утилизации (использования) 1) патология печени из-за снижения ГНГ
(п/л печени – из-за алкоголизма и др.),
2) дефицит В1, т.к. без него не может работать ПДГ и ЦТК.
Превращение лактата в пируват уравнение
а) Анаэробные процессы высвобождения энергии. Анаэробный гликолиз. Иногда в случае отсутствия или недостаточного количества кислорода окислительное фосфорилирование становится невозможным. Но даже при таких условиях небольшое количество энергии может быть доставлено клеткам путем гликолитического расщепления углеводов, поскольку для химической реакции расщепления глюкозы до пировиноградной кислоты кислорода не требуется.
Это чрезвычайно неэкономичный путь метаболизма глюкозы, т.к. только 24000 калорий энергии, выделяемой при расщеплении каждой молекулы глюкозы, используется для образования АТФ, что составляет чуть больше 3% общего количества выделившейся энергии. Тем не менее, такой путь метаболизма, названный анаэробным энергообеспечением, в ситуации, когда кислород недоступен, доставляет энергию в течение нескольких минут, что может оказаться спасительным для клеток.
б) Образование молочной кислоты во время анаэробного гликолиза способствует выделению дополнительного количества энергии сверх анаэробного энергообеспечения. Согласно закону действующих масс, если количество образующихся конечных продуктов реакции приближается к средним значениям, обеспечиваемым реакцией, скорость реакций снижается практически до нуля. Конечными продуктами реакции гликолиза являются (для облегчения понимания просим вас изучить рисунок ниже):
(1) пировиноградная кислота;
(2) атомы водорода, присоединяющиеся к НАД+, образуя НАД-Н и Н+.
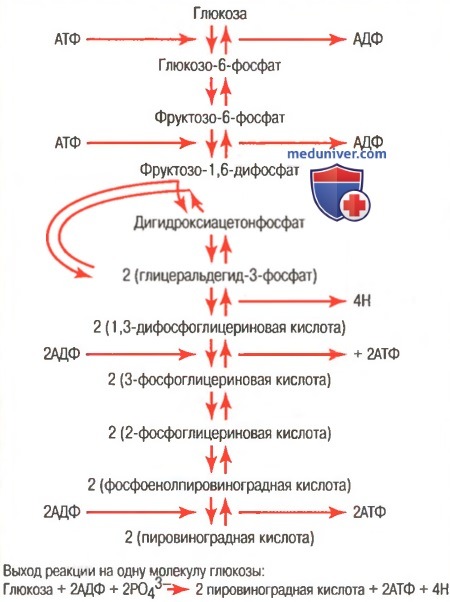 Последовательность химических реакций, ответственных за гликолиз
Последовательность химических реакций, ответственных за гликолиз
Образование обоих или одного из них останавливает процессы гликолиза и препятствует дальнейшему образованию АТФ. Если количество образовавшихся конечных продуктов реакции велико, они взаимодействуют друг с другом, образуя молочную кислоту в соответствии со следующей схемой реакции:
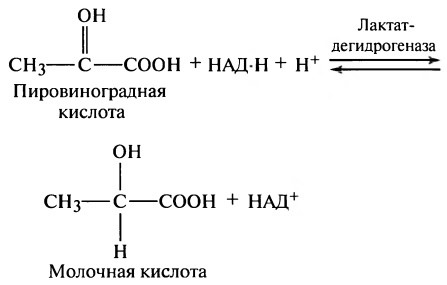
Таким образом, в анаэробных условиях большое количество пировиноградной кислоты превращается в молочную кислоту, которая легко диффундирует во внеклеточное пространство и даже внутрь некоторых менее активных клеток. Следовательно, молочная кислота представляет собой разновидность «водосточного колодца», в котором исчезают конечные продукты гликолиза, что позволяет гликолизу осуществляться дольше, чем это могло быть при отсутствии молочной кислоты.
Без этого превращения пировиноградной кислоты гликолиз мог бы осуществляться лишь в течение нескольких секунд. Вместо этого гликолиз продолжается в течение нескольких минут, снабжая организм значительным дополнительным количеством АТФ даже при отсутствии кислорода.
в) Обратное превращение молочной кислоты в пировиноградную, когда кислород становится доступным. Если кислород вновь становится доступным после периода анаэробного метаболизма, молочная кислота быстро превращается в пировиноградную кислоту, НАД-Н и Н+. Большие количества этих веществ немедленно окисляются, образуя значительное количество АТФ. Избыток АТФ может явиться причиной того, что более 75% пировиноградной кислоты вновь превращается в глюкозу.
Таким образом, большое количество молочной кислоты, которое образуется во время анаэробного гликолиза, не теряется организмом, т.к. если кислород вновь становится доступным, молочная кислота может подвергнуться обратному превращению в глюкозу или напрямую использоваться для получения энергии. Большая часть этих превращений осуществляется в печени, но в небольших количествах может происходить и в других тканях.
г) Использование сердцем молочной кислоты для получения энергии. Сердечная мышца обладает способностью превращать молочную кислоту в пировиноградную и затем использовать последнюю для получения энергии. В большинстве случаев это происходит при больших физических нагрузках, когда в кровь поступают значительное количество молочной кислоты из скелетных мышц и суммарно дает существенное количество энергии сердечной мышце.
Видео этапы, реакции гликолиза
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
БИОХИМИЯ УЧЕБНИК ДЛЯ ВУЗОВ — Е. С. Северина — 2004
РАЗДЕЛ 7. ОБМЕН УГЛЕВОДОВ
IX. Ситез глюкозы в печени (глюконеогенез)
Некоторые ткани, например, мозг, нуждаются в постоянном поступлении глюкозы. Когда поступление углеводов в составе пищи недостаточно, содержание глюкозы в крови некоторое время поддерживается в пределах нормы за счёт расщепления гликогена в печени. Однако запасы гликогена в печени невелики. Они значительно уменьшаются к 6 —10 ч голодания и практически полностью исчерпываются после суточного голодания. В этом случае в печени начинается синтез глюкозы de novo — глюконеогенез.Глюконеогенез — процесс синтеза глюкозы из веществ неуглеводной природы. Его основной функцией является поддержание уровня глюкозы в крови в период длительного голодания и интенсивных физических нагрузок. Процесс протекает в основном в печени и менее интенсивно в корковом веществе почек, а также в слизистой оболочке кишечника. Эти ткани могут обеспечивать синтез 80 — 100 г глюкозы в сутки. На долю мозга при голодании приходится большая часть потребности организма в глюкозе. Это объясняется тем, что клетки мозга не способны, в отличие от других тканей, обеспечивать потребности в энергии за счёт окисления жирных кислот (см. раздел 8).
Кроме мозга, в глюкозе нуждаются ткани и клетки, в которых аэробный путь распада невозможен или ограничен, например, эритроциты (они лишены митохондрий), клетки сетчатки, мозгового слоя надпочечников и др.
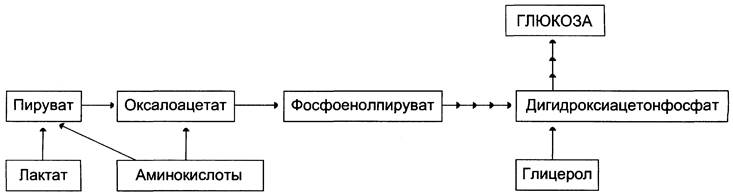
Первичные субстраты глюконеогенеза — лактат, аминокислоты и глицерол. Включение этих субстратов в глюконеогенез зависит от физиологического состояния организма.
✵ Лактат — продукт анаэробного гликолиза. Он образуется при любых состояниях организма в эритроцитах и работающих мышцах. Таким образом, лактат используется в глюконеогенезе постоянно.
✵ Глицерол высвобождается при гидролизе жиров в жировой ткани в период голодания или при длительной физической нагрузке.
✵ Аминокислоты образуются в результате распада мышечных белков и включаются в глюконеогенез при длительном голодании или продолжительной мышечной работе.
На рисунке 7-44 показаны пункты включения первичных субстратов в глюконеогенез.
Рис. 7-44. Включение субстратов в глюконеогенез.
А. Реакции глюконеогенеза
Большинство реакций глюконеогенеза протекает за счёт обратимых реакций гликолиза (рис. 7-45, реакции 9, 8, 7, 6, 5, 4, 2) и катализируется теми же ферментами. Однако 3 реакции гликолиза термодинамически необратимы. На этих стадиях реакции глюконеогенеза протекают другими путями.
Рис. 7-45. Гликолиз и глюконеогенез. Ферменты обратимых реакций гликолиза и глюконеогенеза: 2 — фосфоглюкоизомераза; 4 — альдолаза; 5 — триозофосфатизомераза; 6 — глицеральдегидфосфатдегидрогеназа; 7 — фосфоглицераткиназа; 8 — фосфоглицератмутаза; 9 — енолаза. Ферменты необратимых реакций глюконеогенеза: 11 — пируваткарбоксилаза; 12 — фосфоенолпируваткарбоксикиназа; 13 — фруктозо- 1,6-бисфосфатаза; 14 — глюкозо-6-фосфатаза. I-III —субстратные циклы.
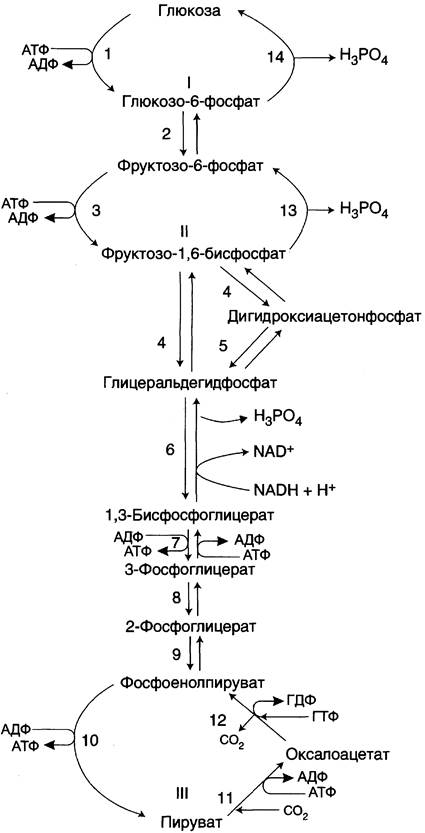
Необходимо отметить, что гликолиз протекает в цитозоле, а часть реакций глюконеогенеза происходит в митохондриях.
Рассмотрим более подробно те реакции глюконеогенеза, которые отличаются от реакций гликолиза и происходят в глюконеогенезе с использованием других ферментов. Рассмотрим процесс синтеза глюкозы из пирувата.
1. Образование фосфоенолпирувата из пирувата — первая из необратимых стадий глюконеогенеза
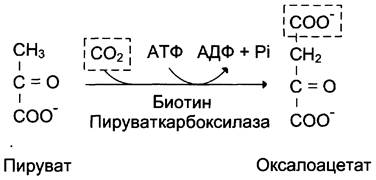
Образование фосфоенолпирувата из пирувата происходит в ходе двух реакций (рис. 7-45, реакции 11, 12), первая из которых протекает в митохондриях. Пируват, образующийся из лактата или из некоторых аминокислот, транспортируется в матрикс митохондрий и там карбоксилируется с образованием оксалоацетата (рис. 7-46). Пируват- карбоксилаза,катализирующая данную реакцию, — митохондриальный фермент, коферментом которого является биотин. Реакция протекает с использованием АТФ.
Рис. 7-46. Образование оксалоацетата из пирувата.
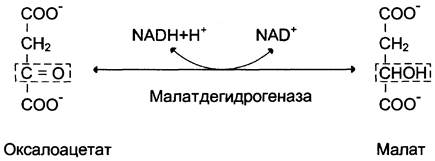
Дальнейшие превращения оксалоацетата протекают в цитозоле. Следовательно, на этом этапе должна существовать система транспорта оксалоацетата через митохондриальную мембрану, которая для него непроницаема. Оксалоацетат в митохондриальном матриксе восстанавливается с образованием малата (рис. 7-47) при участии НАDН (обратная реакция цитратного цикла). Образовавшийся малат затем проходит через митохондриальную мембрану с помощью специальных переносчиков. Кроме того, оксалоацетат способен транспортироваться из митохондрий в цитозоль в виде аспартата в ходе малат-аспартатного челночного механизма, рассмотренного ранее (рис. 7-39).
Рис. 7-47. Превращение оксалоацетата в малат.
В цитозоле малат вновь превращается в оксалоацетат в ходе реакции окисления с участием кофермента NAD + . Обе реакции: восстановление оксалоацетата и окисление малата катализируют малатдегидрогеназа, но в первом случае это митохондриальный фермент, а во втором — цитозольный. Образованный в цитозоле из малата оксалоацетат затем превращается в фосфоенолпируват в ходе реакции, катализируемой фосфоенолпируваткарбоксикиназой — ГТФ-зависимым ферментом (рис. 7-48). Название фермента дано по обратной реакции.
Рис. 7-48. Превращение оксалоацетата в фосфоенолпируват.
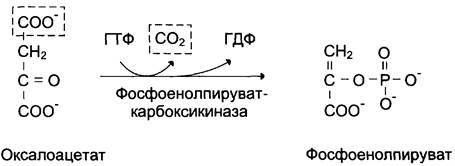
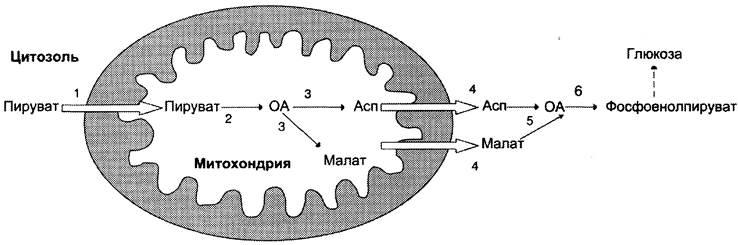
Схема всех реакций, протекающих на первой необратимой стадии глюконеогенеза, представлена на рис. 7-49.
Рис. 7-49. Образование оксалоацетата, транспорт в цитозоль и превращение в фосфоенолпируват. 1 — транспорт пирувата из цитозоля в митохондрию; 2 — превращение пирувата в оксалоацетат (ОА); 3 — превращение ОА в малат или аспартат; 4 — транспорт аспартата и маната из митохондрии в цитозоль; 5 — превращение аспартата и маната в ОА; 6 — превращение ОА в фосфоенолпируват.
Следует отметить, что этот обходной участок глюконеогенеза требует расхода двух молекул с макроэргическими связями (АТФ и ГТФ) в расчёте на одну молекулу исходного вещества — пирувата. В пересчёте на синтез одной молекулы глюкозы из двух молекул пирувата расход составляет 2 моль АТФ и 2 моль ГТФ или 4 моль АТФ (для удобства рассуждений предлагается считать, что энергозатраты на синтез АТФ и ГТФ равны). После образования фосфоенолпирувата все остальные реакции также
протекают в цитозоле вплоть до образования фруктозо-1,6-бисфосфата и катализируются гликолитическими ферментами.
2. Гидролиз фруктозо-1,6-бисфосфата и глюкозо-6-фосфата
Отщепление фосфатной группы из фруктозо-1,6-бисфосфата и глюкозо-6-фосфата — также необратимые реакции глюконеогенеза. В ходе гликолиза эти реакции катализируют специфические киназы с использованием энергии АТФ. В глюконеогенезе они протекают без участия АТФ и АДФ и ускоряются не киназами, а фосфатазами — ферментами, принадлежащими к классу гидролаз. Ферменты фруктозо-1,6-бисфосфатаза и глюкозо-6-фосфатаза катализируют отщепление фосфатной группы от фруктозо-1,6-бисфосфата и глюкозо-6-фосфата. После чего свободная глюкоза выходит из клетки в кровь. Схема всех реакций глюконеогенеза представлена на рис. 7-45.
Итак, в печени существуют 4 фермента, которые принимают участие только в глюконеогенезе и катализируют обходные реакции необратимых стадий гликолиза. Это — пируват- карбоксилаза, фосфоенолпируваткарбоксикиназа, фруктозо-1,6-бисфосфатаза и глюкозо-6- фосфатаза.
3. Энергетический баланс глюконеогенеза из пирувата
В ходе этого процесса расходуются 6 моль АТФ на синтез 1 моль глюкозы из 2 моль пирувата. Четыре моль АТФ расходуются на стадии синтеза фосфоенолпирувата из оксалоацетата и ещё 2 моль АТФ на стадиях образования 1,3-бисфосфоглицерата из 3-фосфоглицерата.
Суммарный результат глюконеогенеза из пирувата выражается следующим уравнением:
2 Пируват + 4 АТФ + 2 ГТФ + 2 (NАDН + Н + ) + 4 Н2O —> Глюкоза + 4 АДФ + 2 ГДФ + 6 Н3РO4 + 2 NАD + .
Б. Синтез глюкозы из лактата
Лактат, образованный в анаэробном гликолизе, не является конечным продуктом метаболизма. Использование лактата связано с его превращением в печени в пируват. Лактат как источник пирувата важен не столько при голодании, сколько при нормальной жизнедеятельности организма. Его превращение в пируват и дальнейшее использование последнего являются способом утилизации лактата.
Лактат, образовавшийся в интенсивно работающих мышцах или в клетках с преобладающим анаэробным способом катаболизма глюкозы, поступает в кровь, а затем в печень. В печени отношение NАDН/NАD + ниже, чем в сокращающейся мышце, поэтому лакгатдегидрогеназная реакция протекает в обратном направлении, т. е. в сторону образования пирувата из лактата. Далее пируват включается в глюконеогенез, а образовавшаяся глюкоза поступает в кровь и поглощается скелетными мышцами. Эту
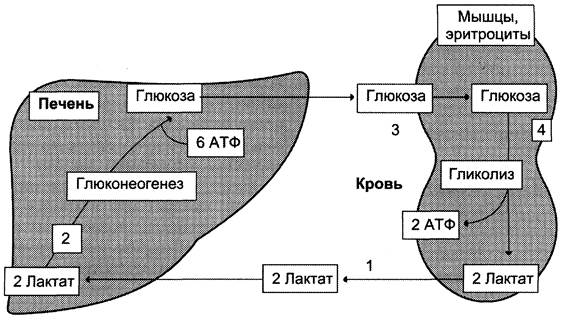
последовательность событий называют «глюкозо-лактатным циклом», или «циклом Кори» (рис. 7-50). Цикл Кори выполняет 2 важнейшие функции: 1 — обеспечивает утилизацию лактата; 2 — предотвращает накопление лактата и, как следствие этого, опасное снижение pH (лактоацидоз). Часть пирувата, образованного из лактата, окисляется печенью до СO2 и Н2O. Энергия окисления может использоваться для синтеза АТФ, необходимого для реакций глюконеогенеза.
Рис. 7-50. Цикл Кори (глюкозо-лактатный цикл). 1 — поступление лактата из сокращающейся мышцы с током крови в печень; 2 — синтез глюкозы из лактата в печени; 3 — поступление глюкозы из печени с током крови в работающую мышцу; 4 — использование глюкозы как энергетического субстрата сокращающейся мышцей и образование лактата.
Лактоацидоз. Термин «ацидоз» обозначает увеличение кислотности среды организма (снижение pH) до значений, выходящих за пределы нормы. При ацидозе либо увеличивается продукция протонов, либо происходит снижение их экскреции (в некоторых случаях и то и другое). Метаболический ацидоз возникает при увеличении концентрации промежуточных продуктов обмена (кислотного характера) вследствие увеличения их синтеза или уменьшения скорости распада или выведения. При нарушении кислотно-основного состояния организма быстро включаются буферные системы компенсации (через 10 — 15 мин). Лёгочная компенсация обеспечивает стабилизацию соотношения НСO3 — /Н2СO3, которая в норме соответствует 1:20, а при ацидозе уменьшается. Лёгочная компенсация достигается увеличением объёма вен
тиляции и, следовательно, ускорением выведения СO2 из организма. Однако основную роль в компенсации ацидоза играют почечные механизмы с участием аммиачного буфера (см. раздел 9). Одной из причин метаболического ацидоза может быть накопление молочной кислоты. В норме лактат в печени превращается обратно в глюкозу путём глюконеогенеза либо окисляется. Кроме печени, другим потребителем лактата служат почки и сердечная мышца, где лактат может окисляться до СO2 и Н2O и использоваться как источник энергии, особенно при физической работе.
Уровень лактата в крови — результат равновесия между процессами его образования и утилизации. Кратковременный компенсированный лактоацидоз встречается довольно часто даже у здоровых людей при интенсивной мышечной работе. У нетренированных людей лактоацидоз при физической работе возникает как следствие относительного недостатка кислорода в мышцах и развивается достаточно быстро. Компенсация осуществляется путём гипервентиляции.
При некомпенсированном лактоацидозе содержание лактата в крови увеличивается до 5 ммоль/л (в норме до 2 ммоль/л). При этом pH крови может составлять 7,25 и менее (в норме 7,36-7,44).
Повышение содержания лактата в крови может быть следствием нарушения метаболизма пирувата (рис. 7-51).
Рис. 7-51. Нарушения метаболизма пирувата при лактоацидозе. 1 — нарушение использования пирувата в глюконеогенезе; 2 — нарушение окисления пирувата.
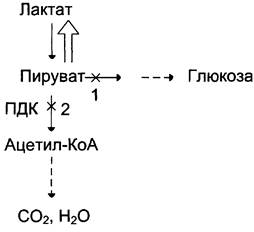
Так, при гипоксии, возникающей вследствие нарушения снабжения тканей кислородом или кровью, уменьшается активность пируватдегидрогеназного комплекса и снижается окислительное декарбоксилирование пирувата. В этих условиях равновесие реакции пируват лактат сдвинуто в сторону образования лактата. Кроме того, при гипоксии уменьшается синтез АТФ, что, следовательно, ведёт к снижению скорости глюконеогенеза — другого пути утилизации лактата. Повышение концентрации лактата и снижение внутриклеточного pH отрицательно влияют на активность всех ферментов, в том числе и пируваткарбоксилазы, катализирующей начальную реакцию глюконеогенеза.
Возникновению лактоацидоза также способствуют нарушения глюконеогенеза при печёночной недостаточности различного происхождения. Кроме того, лактоацидозом может сопровождаться гиповитаминоз В1, так как производное этого витамина (тиаминдифосфат) выполняет коферментную функцию в составе ПДК при окислительном декарбоксилировании пирувата (см. раздел 6). Дефицит тиамина может возникать, например, у алкоголиков с нарушенным режимом питания.
Итак, причинами накопления молочной кислоты и развития лактоацидоза могут быть:
✵ активация анаэробного гликолиза вследствие тканевой гипоксии различного происхождения;
✵ поражения печени (токсические дистрофии, цирроз и др.);
✵ нарушение использования лактата вследствие наследственных дефектов ферментов глюконеогенеза, недостаточности глюкозо-6-фосфатазы;
✵ нарушение работы ПДК вследствие дефектов ферментов или гиповитаминозов;
✵ применение ряда лекарственных препаратов, например, бигуанидов (блокаторы глюконеогенеза, используемые при лечении сахарного диабета).
В. Синтез глюкозы из аминокислот
В условиях голодания часть белков мышечной ткани распадается до аминокислот, которые далее включаются в процесс катаболизма. Аминокислоты, которые при катаболизме превращаются в пируват или метаболиты нитратного цикла, могут рассматриваться как потенциальные предшественники глюкозы и гликогена и носят название гликогенных. Например, оксалоацетат, образующийся из аспарагиновой кислоты, является промежуточным продуктом как нитратного цикла, так и глюконеогенеза.
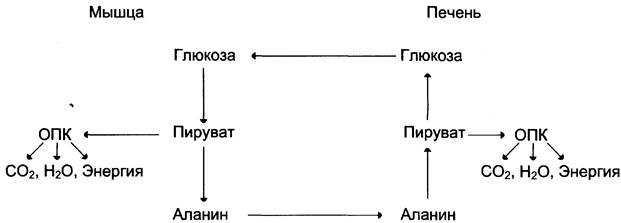
Из всех аминокислот, поступающих в печень, примерно 30% приходится на долю аланина. Это объясняется тем, что при расщеплении мышечных белков образуются аминокислоты, многие из которых превращаются сразу в пируват или сначала в оксалоацетат, а затем в пируват. Последний превращается в аланин, приобретая аминогруппу от других аминокислот. Аланин из мышц переносится кровью в печень, где снова преобразуется в пируват, который частично окисляется и частично включается в глюконеогенез. Следовательно, существует следующая последовательность событий (глюкозо-аланиновый цикл): глюкоза в мышцах —> пируват в мышцах —> аланин в мышцах —> аланин в печени —> глюкоза в печени —> глюкоза в мышцах (рис. 7-52). Весь цикл не приводит к увеличению количества глюкозы в мышцах, но он решает проблемы транспорта аминного азота из мышц в печень и предотвращает лактоацидоз.
Рис. 7-52. Глюкозо-аланиновый цикл.
Г. Синтез глюкозы из глицерола
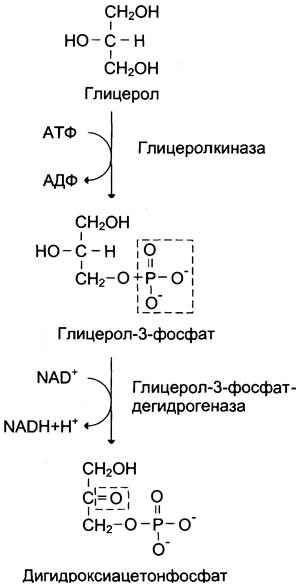
Глицерол образуется при гидролизе триацил- глицеролов, главным образом в жировой ткани. Использовать его могут только те ткани, в которых имеется фермент глицерол киназа, например, печень, почки. Этот АТФ-зависимый фермент катализирует превращение глицерола в α-глицерофосфат (глицерол-3-фосфат). При включении глицерол-3-фосфата в глюконеогенез происходит его дегидрирование NAD-зaвисимой дегидрогеназой с образованием дигидроксиацетонфосфата (рис. 7-53), который далее превращается в глюкозу.
Рис. 7-53. Превращение глицерола в дигидроксиацетон- фосфат.
Биологическая библиотека — материалы для студентов, учителей, учеников и их родителей.
Наш сайт не претендует на авторство размещенных материалов. Мы только конвертируем в удобный формат материалы, которые находятся в открытом доступе и присланные нашими посетителями.
Если вы являетесь обладателем авторского права на любой размещенный у нас материал и намерены удалить его или получить ссылки на место коммерческого размещения материалов, обратитесь для согласования к администратору сайта.
Разрешается копировать материалы с обязательной гипертекстовой ссылкой на сайт, будьте благодарными мы затратили много усилий чтобы привести информацию в удобный вид.
© 2018-2022 Все права на дизайн сайта принадлежат С.Є.А.
http://meduniver.com/Medical/Physiology/1179.html
http://lifelib.info/biochemistry/biochemistry_4/46.html