Дегидрогеназы – ферменты класса оксидоредуктаз (пиридинзависимые, флавинзависимые, аэробные и анаэробные типы, физиология, биохимия)

Дегидрогеназы представляют собой энзимы класса оксидоредуктаз, которые катализируют реакции отщепления водорода (то есть протонов и электронов) от субстрата, который является окислителем, и транспортируют его на другой субстрат, который восстанавливается.
В зависимости от химической природы акцептора, с которым взаимодействуют дегидрогеназы, их делят на несколько групп:
- Анаэробные дегидрогеназы, которые катализируют реакции, в которых акцептором водорода является соединение, которое отличается от кислорода.
- Аэробные дегидрогеназы, которые катализируют реакции где акцептором водорода может быть кислород (оксидазы) или другой акцептор. Аэробные дегидрогеназы относятся к флавопротеинам, продукт реакции — перекись водорода.
- Дегидрогеназы, которые обеспечивают транспортировку электронов от субстрата к акцептору электронов. К этой группе дегидрогеназ принадлежат цитохромы дыхательной цепи митохондрий.
- Дегидрогеназы, которые катализируют прямое введение в молекулу субстрата окисляется, 1 или 2 атомов кислорода. Такие дегидрогеназы получили название оксигеназы.
Функцию первичных акцепторов атомов водорода, отщепляющихся от соответствующих субстратов, выполняют дегидрогеназы 2 типов:
- пиридинзависимые дегидрогеназы — содержат коферменты никотинамид (НАД +) или никотинамидадениндинуклеотидфосфат (НАДФ +).
- флавинзависимые дегидрогеназы, простетической группой которых является флавинадениндинуклеотид (ФАД) или флавинмононуклеотид (ФМН).
Коферменты НАДФ+ (или НАД+) с апоферментом связаны непрочно и поэтому могут в клетке находиться как в связанном с апоферментом состоянии, так и быть отделенными от белковой части.
Пиридинзависимые дегидрогеназы относятся к анаэробному типу — водорастворимые ферменты, окисляющие полярные субстраты. Реакции, катализируемой пиридинзависимыми дегидрогеназами, в общем виде приведены в таких уравнениях:
SH2 + НАДФ+ → S + НАДФН + Н+
SH2 + НАД+ → S + НАДН + Н+
Рабочей структурой в молекуле НАД + или НАДФ + является пиридиновое кольцо никотинамид, который присоединяет в ходе ферментативной реакции один атом водорода и один электрон (гидрид-ион), а второй протон поступает в реакционную среду. Пиридинзависимые дегидрогеназы очень распространены в живых клетках. Они отщепляющие протоны и электроны от многих субстратов, восстанавливая НАД + или НАДФ + и передавая в дальнейшем восстановительные эквиваленты на другие акцепторы. НАД-зависимые дегидрогеназы главным образом катализируют окислительно-восстановительные реакции окислительных путей метаболизма — гликолиза, цикла Кребса, β-окисления жирных кислот, дыхательной цепи митохондрий и др. НАД является главным источником электронов для цепи переноса электронов. НАДФ используется главным образом в процессах восстановительного синтеза (в синтезе жирных кислот и стероидов).
Флавинзависимые дегидрогеназы — флавопротеины, простетическими группами в которых ФАД или ФМН — производные витамина В2, которые прочно (ковалентно) связаны с апоферментом. Данные дегидрогеназы — мембраносвязанные ферменты, окисляющие неполярные и малополярные субстраты. Рабочей частью молекулы ФАД или ФМН, которая участвует в окислительно-восстановительных реакциях, является изоалоксазиновое кольцо рибофлавина, который акцептирует два атома водорода (2Н+ + 2е-) от субстрата.
Общее уравнение реакции с участием флавинзависимых дегидрогеназ выглядит так:
SH2 + ФМН → S + ФМН-Н2
SH2 + ФАД+ → S + ФАДН2
В процессах биологического окисления данные ферменты играют роль как анаэробных, так и аэробных дегидрогеназ. К, анаэробным дегидрогеназам принадлежит НАДН-дегидрогеназа, ФМН-зависимый фермент, который передает электроны от НАДН на более электроположительны компоненты дыхательной цепи митохондрий. Другие дегидрогеназы (ФАД-зависимые) переносят электроны непосредственно от субстрата на дыхательную цепь (например, сукцинатдегидрогеназа, ацил-КоА-дегидрогеназа). Транспортировка электронов от флавопротеинов к цитохромоксидазам в дыхательной цепи обеспечивают цитохромы, которые, кроме цитохромоксидазы, по классификации как анаэробные дегидрогеназы. Цитохромы – представляют собой железосодержащие протеины митохондрий — гемпротеины, которые за счет обратного изменения валентности геминового железа выполняют функцию транспортировки электронов в аэробных клетках непосредственно в цепях биологического окисления: цитохром (Fe3 +) + е → цитохром (Fe2 +).
В состав дыхательной цепи митохондрий входят цитохромы b, с1, с, а и а3 (цитохромоксидаза). Кроме дыхательной цепи, цитохромы содержатся в эндоплазматическом ретикулуме (450 и b5). К аэробным флавинзависимым дегидрогеназам принадлежат оксидазы L-аминокислот, ксантиноксидаза и др.
Дегидрогеназы, катализируют включение одного или двух атомов кислорода в молекулу субстрата, получили название оксигеназы. В зависимости от количества атомов кислорода, которые взаимодействуют с субстратом, оксигеназы делят на 2 группы:
Диоксигеназы катализируют присоединяет в молекулу субстрата 2 атомов кислорода: S + O2 → SO2. Это, в частности, негемовые железосодержащие ферменты, катализирующие реакции синтеза гомогентизиновой кислоты и ее окисления на пути катаболизма тирозина. Железосодержащая липооксигеназа катализирует включение О2 в арахидоновую кислоту, первую реакцию в процессе синтеза лейкотриенов. Пролин- и лизиндиоксигеназы катализируют реакции гидроксилирования остатков лизина и пролина в проколлагене. Монооксигеназы катализируют присоединение к субстрату только 1 из атомов молекулы кислорода. При этом, второй атом кислорода восстанавливается до воды:
SH + О2 + НАДФН + Н+
К монооксигеназам принадлежат ферменты, которые участвуют в обмене веществ многих лекарственных субстанций путем их гидроксилирования. Эти ферменты имеют локализацию преимущественно в микросомальной фракции печени, надпочечников, половых желез и других тканей. Поскольку чаще всего субстрат в монооксигеназных реакциях гидроксилируется, данную группу энзимов также называют гидроксилазы.
Монооксигеназы катализируют реакции гидроксилирования холестерина (стероидов) и превращение их в биологически активные субстанции, в том числе — половые гормоны, гормоны надпочечников, активные метаболиты витамина D — кальцитриол, а также реакции детоксикации путем гидроксилирования ряда токсичных веществ, лекарственных средств и продуктов их превращения для организма. Монооксигеназная мембранная система эндоплазматического ретикулума гепатоцитов содержит НАДФН + Н+, флавопротеины с кофактором ФАД, протеин (адренотоксин), содержащий негемовое железо, и гемпротеин — цитохром Р450. В результате гидроксилирования неполярных гидрофобных субстанций повышается их гидрофильность, что способствует инактивации биологически активных веществ или обезвреживанию токсичных субстанций и экскреции их из организма. Некоторые лекарственные субстанции, такие, как фенобарбитал, имеют способность индуцировать синтез микросомальных ферментов и цитохрома Р450.
Существуют монооксигеназы, которые не содержат цитохрома Р450. К ним относятся ферменты печени, которые катализируют реакции гидроксилирования фенилаланина, тирозина, триптофана.
Полезно знать
© VetConsult+, 2015. Все права защищены. Использование любых материалов, размещённых на сайте, разрешается при условии ссылки на ресурс. При копировании либо частичном использовании материалов со страниц сайта обязательно размещать прямую открытую для поисковых систем гиперссылку, расположенную в подзаголовке или в первом абзаце статьи.
Параграф 20. 2. Виды оксидоредуктаз
Автор текста – Анисимова Елена Сергеевна.
Авторские права защищены. Продавать текст нельзя.
Курсив не зубрить.
Замечания можно присылать по почте: exam_bch@mail.ru
https://vk.com/bch_5
Параграф учебника по биохимии 20.2
«Виды оксидоредуктаз».
см. п.3.
См. таблицу в файле «20.2 Таблица оксидоредуктазы».
Определение.
Оксидоредуктазы – это класс ферментов, которые катализируют окислительно-восстановительные реакции.
Основные группы (не только подклассы) оксидоредуктаз:
дегидрогеназы, оксидазы, оксигеназы,
редуктазы, цитохромы и антиокислительные ферменты.
Что такое окисление и восстановление.
Окисление – это отщепление электрона,
а восстановление – это присоединение электрона.
Окисление и восстановления всегда сопряжены, то есть связаны,
потому что если одно вещество теряет электрон
и при этом окисляется,
то другое вещество этот электрон приобретает
и при этом восстанавливается.
При восстановлении органических субстратов
электроны часто присоединяются в составе атомов водорода
(каждый из которых состоит из одного электрона и одного протона),
источником которых является НАДФН,
который получает атомы водорода в ПФП (п.35).
При окислении органических субстратов
электроны отщепляются обычно в составе атомов водорода,
которые переносятся на кофермент (НАД и др.) или кислород.
Присоединение атомов кислорода
тоже является окислением органического субстрата,
поскольку присоединившиеся атомы кислорода притягивают к себе электронную плотность тех атомов, в которым они присоединились.
1. Д Е Г И Д Р О Г Е Н А З Ы
катализируют реакции дегидрирования,
то есть отщепления атомов водорода (Н).
При этом водород
(2 атома водорода, состоящие из 2 протонов и 2 электронов)
отщепляется от органического субстрата (R)
и переносится на кофермент (на КоФ – НАД+, ФАД или НАДФ+).
Формулируя короче – дегидрогеназы переносят 2 атома водорода
от субстрата на кофермент.
Субстрат до отщепления от него пары атомов водорода
называется восстановленным субстратом
и обозначается так – RН2;
где Н – это атомы водорода, которые будут отщепляться,
а R – это остальные атомы молекулы субстрата.
Субстрат после отщепления от него пары атомов водорода
обозначается так – R
и называется окисленным.
Кофермент, получающий атомы водорода,
до их присоединения обозначается как КоФ,
а после присоединения двух Н обозначается как КоФН2.
Поскольку при отщеплении атомов водорода
вещество теряет вместе с водородом
входящие в состав водорода электроны,
а потеря электронов называется окислением,
то RН2 – более восстановленное вещество, чем R,
а КоФН2 – более восстановленное вещество, чем КоФ.
Поэтому RН2 называют восстановленным субстратом,
а R – окисленным субстратом.
КоФ2 называют восстановленным коферментом
(восстановленной формой кофермента),
а КоФ – окисленным коферментом
(окисленной формой кофермента).
Схема реакции, которая катализируется дегидрогеназами,
может быть представлена в таком виде:
RH2 + КоФ ; R + КоФН2.
Где RH2 – это восстановленный субстрат
(органический субстрат до отщепления водорода),
КоФ – окисленный кофермент
(кофермент до присоединения к нему отщепленного от субстрата водорода),
R – окисленный субстрат
(органический субстрат после отщепления от него водорода),
КоФН2 – восстановленный кофермент
(кофермент после присоединения к нему отщепленного от субстрата водорода).
Коферменты, которые участвуют в работе дегидрогеназ:
НАД+, ФАД и НАДФ+.
НАД+, ФАД и НАДФ+ – это обозначения окисленных форм коферментов,
а НАДН,Н+, ФАДН2 и НАДФН,Н+ – это обозначения восстановленных форм коферментов.
НАДН, Н+ и ФАДН2 переносят водород
от субстратов в ДЦ (для выработки АТФ),
а НАДФН, Н+ переносит водород от метаболитов ПФП не в ДЦ,
а в реакции,
катализируемые редуктазами и гидроксилазами.
Дегидрогеназы катализируют реакции
процессов окислительного катаболизма.
См. п.32, 45, 65.
2. О К С И Д А З Ы
катализируют дегидрирование,
при котором водород, отщепленный от субстрата, присоединяется к кислороду (О2).
Формулируя короче – дегидрогеназы переносят 2 атома водорода
от субстрата на кислород.
Схема реакции, которая катализируется оксидазами,
может быть представлена в таком виде:
RH2 + О2 ; R + Н2О2.
Где RH2 – это восстановленный субстрат,
R – окисленный субстрат
(органический субстрат после отщепления от него водорода).
Кислород в результате этой реакции восстанавливается.
Для работы оксидоредуктаз нужен кофермент ФАД
и витамин В2 в составе ФАД.
Основные примеры оксидаз –
это ксантин/оксидаза (КсО) в п.71
и моноамин/оксидаза (МАО) в п.105.
КсО участвует в катаболизме пуринов
(аденина и гуанина,
но субстраты КсО – гипоксантин и ксантин;
под действием КсО гипоксантин превращается в ксантин,
а ксантин – в мочевую кислоту).
МАО участвует в окислении аминов (адреналина, например).
Ингибиторы КсО применяются для снижения выработки уратов (см. гипер/урикемии),
а ингибиторы МАО применяются в качестве антидепрессантов.
Оксидазы участвуют в реакциях свободного окисления – п.26.
Сходство и отличия дегидрогеназ и оксидаз:
сходство в том, что катализируют отщепление водорода (дегидрирование),
отличия в том, что
дегидрогеназы переносят отщеплённый от субстратов водород на коферменты,
а оксидазы – на кислород (окси).
3. О К С И Г Е Н А З Ы
катализируют включение в молекулу органического субстрата
атомов кислорода, источником которых является О2.
Атомы кислорода включаются
между атомом водорода органического субстрата
и атомом углерода субстрата,
поэтому молекула органического субстрата
до включения в нее атомов кислорода оксигеназами обозначается так – RН,
а после включения атомов кислорода –
либо R-О-О-Н (если включены оба атома О2),
либо R-О-Н (если включен один атом О2).
3.1). Оксигеназы, которые включают в молекулу субстрата
оба атома кислорода,
называются ДИоксигеназами;
схему реакции можно записать так:
Продукт реакции (R-О-О-Н)
называется органической перекисью.
RООН – более окисленное вещество, чем RН,
то есть окисляется органический субстрат,
а восстанавливается кислород.
3.2). Оксигеназы, которые включают в молекулу органического субстрата
только один атом кислорода,
называются МОНОоксигеназами или гидроксилазами.
Второй атом кислорода молекулы кислорода
соединяется с двумя атомами водорода, образуя молекулу воды;
источником атомов водорода в этом случае является кофермент –
либо НАДФН, либо аскорбат.
Схему реакции можно записать так:
RH + О2 + КоФН2 ; R-О-Н + КоФ + Н2О.
RН – это субстрат, в который должен включиться атом кислорода,
R-О-Н – это субстрат со включенным в него атомом кислорода,
О2 – это источник атомов кислорода,
КоФН2 – это источник атомов водорода.
Субстрат RН – более восстановленный по сравнению с RОН.
В реакции окисляются RН и КоФН2,
а восстанавливается кислород.
-ОН – это спиртовая (гидроксильная) группа,
а RОН – это спирт.
Из-за того, что при действии монооксигеназ
образуется гидроксильная группа,
у них есть синоним – гидроксилазы. (Не путать с гидролазами.)
Нужно уметь написать реакции,
которые катализируют фенилаланин/гидроксилаза
и тирозин/гидроксилаза (п.105 и 68).
И знать значение этих реакций.
Оксигеназы (как и оксигеназы),
участвуют в процессах свободного окисления.
Участвуют в метаболизме ксенобиотиков,
в обмене многих гормонов,
в образовании коллагена.
4. Р Е Д У К Т А З Ы
катализируют присоединение водорода
к органическому субстрату,
при этом источником атомов водорода является НАДФН.
Другой вариант определения –
редуктазы катализируют перенос водорода
от кофермента на органический субстрат.
Действие редуктаз противоположно действию дегидрогеназ.
При получении водорода субстрат восстанавливается,
поэтому редуктазы можно считать восстановителями субстратов.
Схема реакций, которые катализируются редуктазами:
R + КоФН2 ; R + КоФН2.
Здесь R – органический субстрат до присоединения водорода,
RН2 – органический субстрат после присоединения к нему атомов водорода,
КоФ2 – восстановленная форма кофермента (источник водорода),
КоФ – окисленная форма кофермента.
В этой реакции органический субстрат восстанавливается (приобретает водород),
а кофермент окисляется (теряет водород).
Редуктазы – основные ферменты восстановительных синтезов –
например, синтеза жирных кислот,
синтеза холестерина (ГМГ-КоА/редуктаза),
синтеза дезокси/рибонуклеотидов (рибонуклеотид/редуктаза).
А глутатион/редуктаза относится к антиокислительным ферментам – п.27.
5. Ц И Т О Х Р О М Ы
участвуют в реакциях изменения валентности металлов –
например, в присоединении электрона к иону железа Fe3+.
В результате ион железа Fe3+превращается в ион железа Fe2+
(восстановление иона железа).
Схема реакции: Fe3+ + е ; Fe2+.
Почему (3+) превращается в (2+) при присоединении электрона – потому что у электрона отрицательный заряд.
Получается (3+) + (1-) = (2+).)
В каких процессах встречаются цитохромы –
цитохромы катализируют ряд реакций дыхательной цепи – п.22;
нужно также помнить цитохром Р 450 – п.118.
6. А Н Т И О К И С Л И Т Е Л Ь Н Ы Е Ф Е Р М Е Н Т Ы .
См. п.27.
Это ферменты, которые участвуют в разрушении активных форм кислорода
и восстановлении окисленных ими веществ клетки
(органических перекисей).
Относятся к антиокислительной системе организма,
к которой также относятся низкомолекулярные антиоксиданты
(витамины А, Е, С и другие).
Благодаря этой системе организм дольше сохраняет молодость
и дольше не появляются такие болезни, как атеросклероз, катаракта и т.д..
Подробнее об антиокислительных ферментах сказано в ответе на вопрос № 27.
Примеры антиокислительных ферментов –
супероксид/дисмутаза,
глутатион/пероксидаза, глутатион/трансфераза,
глутатион/редуктаза, каталаза.
Реакции, катализируемые оксидоредуктазами,
можно разделить на группы –
к одной группе относятся оксидоредуктазы,
которые катализируют восстановление органических субстратов
(редуктазы = восстановители),
а к другой – оксидоредуктазы, которые катализируют
окисление органических субстратов (все, кроме редуктаз).
Оксидоредуктазы, которые катализируют реакции окисления органических субстратов,
можно разделить на три группы:
1) катализирующие отщепление водорода
(дегидрирование) дегидрогеназы и оксидазы,
2) катализирующие присоединение кислорода
(оксигенацию) оксигеназы,
3) катализирующие изменение валентности металлов цитохромы.
Как определить, какая форма субстрата более восстановленная,
а какая – более окисленная:
чем больше водорода в веществе,
тем более восстановлено вещество;
чем больше кислорода в веществе,
тем более окисленное вещество.
Дегидрогеназы, строение и действие
Окисление органических веществ в живых тканях растений и животных происходит с участием ферментов: дегидрогеназ – активаторов и переносчиков водорода (электрона) дыхательного материала и оксидаз – активаторов молекулярного кислорода.
Дегидрогеназы (от де. и новолат. hуdrogenium – водород), ферменты, катализирующие отщепление водорода от органических веществ. Коферментами Д. обычно являются динуклеотиды:
— никотинамидадениндинуклеотидфосфат (НАДФ) или флавинадениндинуклеотид (ФАД)
— флавинмононуклеотид (ФМН), которые акцептируют водород окисляемого вещества. Д. осуществляют первый этап биологического окисления.
Они играют большую роль в цикле Кребса, в гликолизе и в пентозофосфатном цикле. Некоторые Д., не связанные с коферментами, катализируют реакции окисления веществ непосредственно кислородом. Большая часть Д. в составе активного центра содержит металлы – цинк, марганец.
Дегидрогеназы действуют на протяжении всего процесса дыхания и по характеру действия делятся на аэробные и анаэробные. Первые переносят водород непосредственно на молекулярный кислород, а вторые – на какой-либо акцептор водорода. Дегидрогеназы относятся к двухкомпонентным ферментам – протеидам с пиридиновыми или флавиновыми (аллоксазиновыми) коферментами.
Пиридиновые дегидрогеназы относятся к группе анаэробных дегидрогеназ, которые являются акцепторами водорода субстрата. Специфичность их действия заключается в способности гидрирования и дегидрирования пиридиновых ядер.
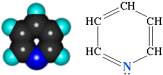
К флавопротеидным дегидрогеназам относятся ферменты, в состав которых фосфорный эфир рибофлавина.
Рис. 2 − Рибофлавин
1. Березов Т. Т., Коровкин Б. Ф.Биологическая химия. М.: Медицина, 1990. С. 213–220; 1998. С. 305–317.
2. Николаев А. Я.Биологическая химия. М.: Высшая школа, 1989. С. 199–221.
3. Филиппович Ю. Б. Основы биохимии. М.: Высшая школа, 1993. С. 403–438.
4. Марри Р. и др. Биохимия человека. М.: Мир, 1993. Т. 1. С. 111–139.
5. Ленинджер А.Основы биохимии. М.: Мир, 1985. Т. 2. С. 403–438, 508–550.
6. АлбертсБ.идр.,Молекулярнаябиология клетки. М.: Мир, 1994.Т. 1. С. 430–459.
7. Скулачев В.П. Энергетика биологическихмембран. М.: Наука. 1989.
8. Это процесс потребление клетками тканей организма кислорода, который участвует в биологическом окислении. Такой вид окисления называют аэробным окислением. Если конечным акцептором в цепи переноса водорода выступает не кислород, а другие вещества (например пировиноградная кислота), то такой тип окисления называют анаэробным.
9. Т.о. биологическое окисление — это дегидрирование субстрата с помощью промежуточных переносчиков водорода и его конечного акцептора. Биологическое окисление питательных веществ происходит в митохондриях. В них были обнаружены ферменты, участвующие в цикле лимонной кислоты, дыхательной цепи, окислительного фосфорилирования, в расщеплении жирных кислот и ряда аминокислот.
10. Дыхательная цепь (ферменты тканевого дыхания) — это переносчики протонов и электронов от окисляемого субстрата на кислород. Окислитель — это соединение, способное принимать электроны. Такая способность количественно характеризуется окислительно-восстановительным потенциалом по отношению к стандартному водородному электроду, рН которого равен 7,0. Чем меньше потенциал соединения, тем сильнее его восстанавливающие свойства и наоборот.
11. Особенности тканевого дыхания
12. Процесс тканевого дыхания нельзя считать тождественным процессам биологического окисления (ферментативным процессам окисления различных субстратов, протекающим в животных, растительных и микробных клетках), поскольку значительная часть таких окислительных превращений в организме происходит в анаэробных условиях, т.е. без участия молекулярного кислорода, в отличие от дыхания тканей.
Большая часть энергии в аэробных клетках образуется благодаря дыханию тканей, и количество образующейся энергии зависит от его интенсивности. Интенсивность Д. т. определяется скоростью поглощения кислорода на единицу массы ткани; в норме она обусловлена потребностью ткани в энергии. Интенсивность его наиболее высока в сетчатке глаза, почках, печени; она значительна в слизистой оболочке кишечника, щитовидной железе, яичках, коре головного мозга, гипофизе, селезенке, костном мозге, легких, плаценте, вилочковой железе, поджелудочной железе, диафрагме, сердце, скелетной мышце, находящейся в состоянии покоя. В коже, роговице и хрусталике глаза интенсивность тканевого дыхания невелика. Гормоны щитовидной железы, жирные кислоты и другие биологически активные вещества способны активизировать тканевое дыхание.
Интенсивность такого дыхания определяют полярографически или манометрическим методом в аппарате Варбурга. В последнем случае для характеристики используют так называемый дыхательный коэффициент — отношение объема выделившегося углекислого газа к объему кислорода, поглощенного определенным количеством исследуемой ткани за определенный промежуток времени.
13. Т. о. любое соединение может отдавать электроны только соединению с более высоким окислительно-восстановительным потенциалом. В дыхательной цепи каждое последующее звено имеет более высокий потенциал, чем предыдущее.
14. Дыхательная цепь
15. Дыхательная цепь состоит из: НАД — зависимой дегидрогеназы; ФАД- зависимой дегидрогеназы; Убихинона (КоQ); Цитохрмов b, c, a+a3 .
16. НАД-зависимые дегидрогеназы. В качестве кофермента содержат НАД и НАДФ. Пиридиновое кольцо никотинамида способно присоединять электроны и протоны водорода.
17. ФАД и ФМН-зависимые дегидрогеназы содержат в качестве кофермента фосфорный эфир витамина В2 (ФАД).
18. Убихинон (КоQ) отнимает водород у флавопротеидов и превращается при этом в гидрохинон.
19. Цитохромы — белки хромопротеиды, способные присоединять электроны, благодаря наличию в своем составе в качестве простетических групп железопорфиринов. Они принимают электрон от вещества, являющегося немного боле сильным восстановителем, и передают его более сильному окислителю. Атом железа связан с атомом азота имидазольного кольца аминоксилоты гистидина с одной стороны от плоскости порфиринового цикла, а с другой стороны с атомом серы метионина. Поэтому потенциальная способность атома железа в цитохромах к связыванию кислорода подавлена.
20. В цитохроме с порфириновая плоскость ковалентно связана с белком через два остатка цистеина, а в цитохромах b и а, она ковалентно не связано с белком.
21. В цитохроме а+а3 (цитохромоксидазе) вместо протопорфирина содержатся порфирин А, который отличатся рядом структурных особенностей. Пятое координационное положение железа занято аминогруппой, принадлежащей остатку аминосахара, входящего в состав самого белка.
22. В отличии от гема гемолгобина атом железа в цитохромах может обратимо переходить из двух в трехвалентное состояние это обеспечивает транспорт электронов.
23. Потребности тканей в кислороде и его запасы
Потребности тканей в кислороде зависят от функционального состояния клеток, входящих в ее состав. Скорость потребления кислорода обычно выражается в мл кислорода на 1 г веса в минуту. В состоянии покоя кислород относительно интенсивно поглощается миокардом, серым веществом головного мозга (в частности, корой головного мозга), печенью и корковым веществом почек. В тоже время скелетные мышцы, селезенка и белое вещество головного мозга в покое потребляют мало кислорода.
При увеличении активности какого-либо органа потребность его в кислороде увеличивается. При физической нагрузке потребление кислорода миокардом может увеличиться в 3 — 4 раза, а работающими скелетными мышцами — более чем в 20 — 50 раз по сравнению с покоем. Потребление кислорода почками возрастает при увеличении интенсивности реабсорбции ионов натрия.
Количество кислорода, которое клетки могут использовать для окислительных процессов, зависит от конвекционного переноса кислорода кровью и диффузии кислорода из капилляров в ткани. Поскольку единственным запасом кислорода в большинстве тканей служит его физически растворенная фракция, снижение поступления кислорода приводит к кислородному голоданию и к замедлению окислительных процессов.
Единственной тканью, в которой имеются запасы кислорода, является мышечная ткань. Роль депо кислорода играет пигмент миоглобин, способный обратимо связывать кислород. Однако содержание миоглобина в мышцах человека невелико, так, среднее содержание миоглобина в сердце составляет 4 мг/г. Поскольку 1 г миоглобина может связать примерно до 1,34 мл кислорода, запасы кислорода в сердце составляют около 0,005 мл кислорода на 1г ткани. Этого количества в условиях полного прекращения доставки кислорода к миокарду может хватить для того, чтобы поддерживать окислительные процессы лишь в течение примерно 3 — 4 секунд.
Миоглобин играет роль кратковременного депо кислорода. В миокарде кислород, связанный с миоглобином, обеспечивает окислительные процессы в тех участках, кровоснабжение которых на короткий срок снижается или полностью прекращается во время систолы.
В начальном периоде интенсивной мышечной нагрузки увеличенные потребности скелетных мышц в кислороде частично удовлетворяются за счет кислорода, высвобождающегося миоглобином. В дальнейшем возрастает мышечный кровоток, и поступление кислорода к мышцам вновь становится адекватным. Восполнение запасов оксимиоглобина является составной частью кислородного долга, который должен быть покрыт каждым мышечным волокном после окончания работы.
Кислородное голодание тканей
При ряде патологических состояний страдает снабжение тканей кислородом. В этих случаях энергетические потребности клеток могут в течение короткого времени удовлетворяться за счет ограниченных запасов энергии в виде АТФ и креатинфосфата, а также за счет анаэробного гликолиза. Однако эти источники энергии недостаточны и могут использоваться недолго, так как в анаэробных условиях резко возрастает потребность клеток в глюкозе, поступление которой обычно не может удовлетворять эту потребность, и во-вторых, в процессе гликолиза образуется большое количество лактата, который медленно удаляется из ткани для последующей переработки (например, для расщепления в печени, почках или миокарде, или для синтеза гликогена). При значительном недостатке кислорода содержание лактата в крови постоянно нарастает, что приводит к нереспираторному ацидозу. Когда рН внутриклеточной среды падает ниже уровня оптимальной активности ферментных систем, наступают выраженные нарушения клеточного метаболизма.
Основные причины, приводящие к кислородному голоданию (тканевой гипоксии), это понижение напряжения кислорода в артериальной крови (артериальная гипоксия), уменьшение кислородной емкости крови (анемия) и уменьшение кровоснабжения того или иного органа (ишемия).
NADН-дегидрогеназа содержит несколько центров типа Fe2S2 и Fe4S4. Атомы железа в таких центрах могут принимать и отдавать электроны поочерёдно, переходя в ферро- (Fe 2 +) и ферри-(Fe 3 +) состояния. От железо-серных центров электроны переносятся на кофермент Q (убихинон) (рис.).
http://proza.ru/2017/06/19/275
http://poisk-ru.ru/s39714t3.html