Механизм возникновения потенциала покоя на биологических мембранах. Формула Нернста, формула Гольдмана
Потенциал покоя – это стационарная разность потенциалов между внешней и внутренней средой клетки, существующая на наружной мембране клетки в невозбужденном состоянии.
Зарегистрировать эту трансмембранную разность потенциалов можно при использовании микроэлектродной техники. Стеклянный микроэлектрод представляет собой стеклянную микропипетку с оттянутым очень тонким кончиком (диаметр кончика 0,1-0,5 мкм), заполненную раствором электролита (обычно 3 М раствором KCl). Таким электродом можно проколоть наружную мембрану клетки, не повредив ее. Второй электрод (электрод сравнения) помещают в раствор у наружной поверхности клетки. Регистрирующее устройство, содержащее усилитель постоянного тока, позволяет измерить трансмембранную разность потенциалов. При этом в нервных и мышечных волокнах различных животных регистрируется разность потенциалов, равная примерно 80-90 мВ (за исключением гладкомышечных клеток, потенциал покоя которых ниже (- 30 мВ), причем внутренняя поверхность клеточной мембраны имеет отрицательный потенциал по отношению к внешней.
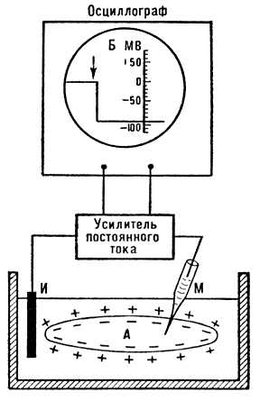
Как же реализуются обязательные условия биоэлектрогенеза на наружной мембране клетки в состоянии покоя?
1) В цитоплазме позвоночных животных преобладают калиевые соли высокомолекулярных соединений (кислот), тогда как в межклеточной среде гораздо выше концентрация натриевых солей неорганических кислот. Например, в гигантском аксоне кальмара (очень удобном для исследований биоэлектрогенеза в связи с большим диаметром) внутриклеточная концентрация К + равна 410 ммоль/л, во внеклеточной среде – 10 ммоль/л. Соответственно, концентрация натрия составляет 49 и 460 ммоль/л. Сходные соотношения наблюдаются и в других клетках.
2) В покое проницаемость наружной мембраны клетки для ионов калия значительно больше, чем для натрия, и больше, чем для ионов хлора:
Например, для аксона кальмара:
Эта проницаемость обусловлена наличием в наружной мембране так называемых потенциалнезависимых каналов, избирательно пропускающих ионы калия, натрия или хлора.
Если концентрация какого-либо иона внутри клетки отлична от концентрации этого иона снаружи и мембрана проницаема для этого иона, возникает поток заряженных частиц через мембрану, вследствие чего электрическая нейтральность системы нарушается, образуется разность потенциалов между наружной и внутренней средой, которая препятствует дальнейшему переносу ионов через мембрану. При установлении равновесия между концентрационным и электрическим градиентом выравниваются значения электрохимических потенциалов по разные стороны мембраны и устанавливается равновесный потенциал для этого иона, который рассчитывается по формуле Нернста.
 μ = μ0 + RTlnC + zFφ
μ = μ0 + RTlnC + zFφ
φ равн. = φi – φe = 
Если принять, что мембранный потенциал обусловлен только переносом ионов калия, то калиевый равновесный потенциал равен:
φ равн. =  -4 % ионов калия от общего их содержания внутри клетки (изменение концентрации калия всего на 2 . 10 -3 ммоль/л). Это пренебрежимое малое количество ионов по сравнению с общим их количеством в клетке.
-4 % ионов калия от общего их содержания внутри клетки (изменение концентрации калия всего на 2 . 10 -3 ммоль/л). Это пренебрежимое малое количество ионов по сравнению с общим их количеством в клетке.
Причина расхождения рассчитанного по формуле Нернста и экспериментального значения мембранного потенциала состоит в том, что не учтена проницаемость мембраны для других ионов (натрия и хлора). Более точно с экспериментальными данными совпадает результат расчета по уравнению Гольдмана:

В числителе выражения, представленного под знаком логарифма, представлены концентрации калия и натрия внутри клетки и концентрация хлора снаружи, в знаменателе – наоборот, это связано с тем, что ионы хлора заряжены отрицательно.
Мембранный потенциал, рассчитанный по уравнению Гольдмана, по абсолютной величине меньше мембранного потенциала, рассчитанного по уравнению Нернста, и ближе к экспериментальным значениям.
И формула Нернста, и формула Гольдмана не учитывают активного транспорта ионов через мембрану – наличия в мембране электрогенного биологического насоса – K + -Na + -АТФазы, перекачивающей калий внутрь клетки, а натрий наружу в неравновесных соотношениях. Чаще всего АТФаза работает в режиме 3Na:2K, то есть за счет работы АТФазы мембранный потенциал больше по абсолютной величине, чем мембранный потенциал, рассчитанный по уравнению Гольдмана.
Если прекратить поступление Na + в клетку, например, путем замещения внеклеточного натрия на такой неспособный к диффузии катион, как холин, то МП будет близок к калиевому равновесному потенциалу.
Нарушение работы K + -Na + -АТФазы приводит к уменьшению мембранного потенциала; в этом случае МП лучше описывается уравнением Гольдмана.
Повреждение клеточной мембраны приводит к повышению проницаемости мембраны для всех ионов. В этом случае разница в проницаемостях сглаживается, МП уменьшается.
Для сильно поврежденных клеток МП еще меньше, но отрицательный мембранный потенциал сохраняется за счет наличия в клетке полианионов – отрицательно заряженных белков, нуклеиновых кислот и других крупных молекул, которые не могут проникнуть через мембрану (доннановский потенциал).
Потенциал покоя. Уравнение Нернста. Уравнение Гольдмана-Ходжкина-Катца
Потенциал покоя (ПП, мембранный потенциал покоя). ПП называется разность потенциалов между внутренней и наружной сторонами клеточной мембраны, возникающей в состоянии покоя клетки. У живых клеток в покое между внутренним содержимым клетки и наружным раствором существует отрицательная разность потенциалов (ПП) порядка 60—90 мВ, которая локализована на поверхностной мембране. Внутренняя сторона мембраны заряжена электроотрицательно по отношению к наружной. ПП обусловлен избирательной проницаемостью покоящейся мембраны для ионов К + (Ю. Бернштейн, 1902, 1912. А. Ходжкин и Б. Катц, 1947).
Предположим, что клетку с мембраной, проницаемой только для ионов калия, поместили в электролит, где их концентрация меньше, чем внутри клетки. Сразу после соприкосновения мембраны с раствором ионы калия начнут выходить из клетки наружу, как выходит газ из надутого шара. Но каждый ион несет с собой положительный электрический заряд, и чем больше ионов калия покинет клетку, тем более электроотрицательным станет ее содержимое. Поэтому на каждый ион калия, выходящий из клетки, будет действовать электрическая сила, препятствующая его движению наружу. В конце концов, установится равновесие, при котором электрическая сила, действующая на ион калия в канале мембраны, будет равна силе, обусловленной различием концентраций ионов калия внутри и вне клетки. Очевидно, что в результате такого равновесия между внутренним и наружным растворами появится разность потенциалов. При этом, если за нуль потенциала принять потенциал внешнего раствора, то потенциал внутри клетки будет отрицательным. Эта разность потенциалов – самое простое из наблюдаемых биоэлектрических явлений – носит название «потенциал покоя» клетки. В первом приближении величину ПП можно рассчитать с помощью формулы Нернста:
 ,
,
где  — концентрация ионов калия внутри клетки,
— концентрация ионов калия внутри клетки,  — концентрация ионов калия снаружи клетки.
— концентрация ионов калия снаружи клетки.
Подставляя численные значения в формулу Нернста, получим значение ПП порядка -87 мВ, что весьма близко к экспериментально измеренному значению ПП. Дальнейшие исследования показали, что вклад в формирование ПП вносят не только ионы калия, но и ионы натрия и хлора.
Концентрация К + в протоплазме примерно в 50 раз выше, чем во внеклеточной жидкости, поэтому, диффундируя из клетки, ионы выносят на наружную сторону мембраны положительные заряды, при этом внутренняя сторона мембраны, практически не проницаемой для крупных органических анионов, приобретает отрицательный потенциал. Поскольку проницаемость мембраны в покое для Na + примерно в 100 раз ниже, чем для К + , диффузия натрия из внеклеточной жидкости (где он является основным катионом) в протоплазму мала и лишь незначительно снижает ПП, обусловленный ионами К + .
В скелетных мышечных волокнах в возникновении потенциала покоя важную роль играют также ионы Cl — , диффундирующие внутрь клетки. Следствием ПП является ток покоя, регистрируемый между поврежденным и интактным участками нерва или мышцы при приложении отводящих электродов. Мембраны нервных и мышечных клеток (волокон) способны изменять ионную проницаемость в ответ на сдвиги мембранного потенциала. При увеличении ПП (гиперполяризация мембраны) проницаемость поверхностных клеточных мембран для Na + и К + падает, а при уменьшении ПП (деполяризация) она возрастает, причём скорость изменений проницаемости для Na + значительно превышает скорость увеличения проницаемости мембраны для К + . Более точные значения для ПП вычисляются по формуле Гольдмана-Ходжкина-Катца (ГХК), которая учитывает проницаемость КМ в покое не только для ионов калия, но и для ионов натрия и хлора:
 ,
,
где PK, PNa, PCl – проницаемость КМ для ионов калия, натрия и хлора, выражения […]i и […]0 обозначают концентрации соответствующих молекул внутри и вне клетки.
В качестве примера вычислим величину ПП для гигантского аксона кальмара. Концентрации ионов внутри и вне аксона приведены ниже (см. таблицу).
| Ион | Концентрация (моль на 1 кг воды) |
| Внутри (i) | Вне (0) |
| Na + | |
| K + | |
| Cl — |
В состоянии покоя при физиологических условиях соотношение коэффициентов проницаемости равно:
Диффузия ионов калия и хлора идет через КМ в обе стороны. Натрий за счет механизма пассивного транспорта проникает внутрь аксона и за счет активного транспорта выносится из клетки. Следовательно, основной вклад в формирование ПП вносят ионы калия и хлора. Подставив численные значения проницаемостей и концентраций для ионов калия хлора в формулу Гольдмана-Ходжкина-Катца для температуры t=30 0 C вычислим:

Значение ПП, вычисленное по формуле Нернста, равно:

Формула Нернста дает несколько заниженное значение потенциала покоя, а формула ГХК приводит к более реалистичным значениям ПП, измеренным экспериментально на крупных клетках.
Следует также отметить, что ни формула Нернста, ни формула ГХК не учитывают механизма активного транспорта.
Формула Томаса для ПП учитывает работу электрогенных ионных Na-K- насосов и имеет вид:
 ,
,
где m – отношение количества ионов натрия к количеству ионов калия, переносимых натрий-калиевым насосом через КМ. Наиболее распространенный режим работы Na + -K + -АТФ-азы наблюдается при m=3/2. В уравнении Томаса отсутствуют члены PCl[Cl — ], так как нет активного транспорта для ионов хлора через КМ.
Численное значение ПП по формуле Томаса равно:
 .
.
Коэффициент m, применяемый в уравнении Томаса, усиливает вклад градиента концентрации ионов калия в формирование ПП. Поэтому ПП, рассчитанный по формуле Томаса, по абсолютному значению меньше ПП, рассчитанного по формуле ГХК. Значение ПП, полученное с помощью формулы Томаса, хорошо совпадает со значениями ПП, измеренными экспериментально на мелких клетках.
Мембранный потенциал покоя
Мембранный потенциал покоя (МПС) — это разность потенциалов между внешней и внутренней сторонами мембраны в условиях, когда клетка не возбуждено. Цитоплазма клетки заряжена отрицательно к внеклеточной жидкости неравномерным распределением анионов и катионов по обе стороны мембраны. Разность потенциалов (напряжение) для различных клеток имеет значение от -50 до -200 мВ (минус означает, что внутри клетка более негативно заряжена, чем снаружи). Мембранный потенциал покоя возникает на мембранах всех клеток — возбуждающих (нервов, мышц, секреторных клеток) и незбудливих.
МПС необходим для поддержания возбудимости таких клеток, как мышечные и нервовои. Также он влияет на транспорт всех заряженных частиц в любом типе клеток: он способствует пассивному транспорта анионов из клетки и катионов в клетку.
Образование и поддержания мембранного потенциала обеспечивают различные типы ионных насосов (в частности натрий-калиевый насос или натрий-калиевая АТФаза) и ионных каналов (калиевые, натриевые, хлорные ионные каналы).
Регистрация потенциала покоя
Для регистрации потенциала покоя используют специальную микроэлектродную технику. Микроэлектрод — это тоненькая стеклянная трубочка, с вытянутым концом, диаметром менее 1 мкм, заполненная раствором электролита (чаще хлорида калия). Рефернтним электродом служит серебряная хлорированная пластинка, расположенная в внеклеточном пространстве, оба электрода подключены к осциллографа. Сначала оба электрода занходяться в внеклеточном пространстве и разность потенциалов между ними отсутствует, если ввести регистрирующий микроэлектрод через мембрану в клетку, то осциллограф покажет скачкообразное смещение потенциала примерно до -80 мВ. Этот сдвиг потенциала называют мембранным потенциалом покоя.
Формирование потенциала покоя
К возникновению мембранного потенциала покоя приводят два фактора: во-первых, концентрации различных ионов отличаются внешне и всереднини клетки, во-вторых мембрана является полупроницаемой: одни ионы могут через нее проникать, другие — нет. Оба эти явления зависят от наличия в мембране специальных белков: концентрационные градиенты создают ионные насосы, а проницаемость мембраны для ионов обеспечивают ионные каналы. Важнейшую роль в формировании мембранного потенциала играют ионы калия, натрия и хлора. Концентрации этих ионов видризняюються по обе стороны мембраны. Для нейрона млекопитающих концентрация K + составляет 140 ммоль внутри клетки и только 5 мМ извне, градиент концентрации Na + почти противоположный — 150 ммоль снаружи и 15 мМ внутри. Такое распределение ионов поддерживается натрий-калиевым насосом в плазматической мембране — белком использующий энергию АТФ для закачки K + в клетку и скачивания Na + из нее. Также существует концентрационный градиент и для других ионов, например, хлорид аниона Cl -.
Концентрационные градиенты катионов калия и натрия — это химическая форма потенциальной энергии. В преобразовании энергии в электрическую участвуют ионные каналы — поры формируются скоплениями специальных трансмембранных белков. Когда ионы диффундируют через канал, они переносят единицу электрического заряда. Любой суммарный движение положительных или отрицательных ионов через мембрану будет создавать напряжение, или разность потенциалов по обе стороны мембраны.
Ионные каналы, участвующие в утовренни МПС имеют избирательную проницаемость, то есть дают возможность проникать только определенному типу ионов. В мембране нейрона в состоянии покоя открытые калиевые каналы (те, что в основном пропускают только калий), большинство натриевых каналов — закрыты. Диффузия ионов K + через калиевые каналы является решающим для создания мембранного потенциала. Так как концентрация K + значительно выше внутри клетки, химический градиент способствует оттоку этих катионов из клетки, поэтому в цитоплазме начинают преобладать анионы, которые не могут проходить через калиевые каналы.
Отток ионов калия из клетки ограничен самым мембранным потенциалом, поскольку при определенном его уровне накопление отрицательных зарядов в цитоплазме будет ограничивать движение катионов за пределы клетки. Таким образом, главным фактором в возникновении МПС является распределение ионов калия под действием электрического и химического потенциалов.
Равновесный потенциал
Для того, чтобы определить влияние движения определенного иона через полупроницаемую мембрану на формирование мембранного потенциала, строят модельные системы. Такая модельная система состоит из сосуда разделенной на две ячейки искусственной полупроницаемой мембраной, в которую встроены ионные каналы. В каждую ячейку можно погрузить электрод и померить разность потенциалов.
Рассмотрим случай, когда искусственная мембрана проницаема только для калия. По две стороны мембраны модельной системы создают градиент концентрации аналогичный таковому у нейроне: в ячейку, соответствующую цитоплазме (внутренняя ячейка), помещают 140 мМ раствор хлорида калия (KCl), в ячейку, соответствующую межклеточной жидкости (внешняя ячейка) — 5 ммоль раствор KCl. Ионы калия будут диффундировать через мембрану во внешнюю ячейку по градиенту концентрации. Но поскольку анионы Cl — проникать через мембрану не могут во внутренней ячейке возникать избыток отрицательного заряда, который будет препятствовать подали оттока катионов. Когда такие модельные нейроны достигнут состояния равновесия, действие химического и электрического потенциала будет сбалансирована, ни суммарной диффузии К + не будет наблюдаться. Значение мембранного потенциала, виинкае при таких условиях, называется равновесным потенциалом для определенного иона (Е ион). Равновесный потенциал для калия составляет примерно -90 мВ.
Аналогичный опыт можно провести и для натрия, установив между ячейками мембрану проникающей только для этого катиона, и поместив во внешнюю ячейку раствор хлорида натрия с концентрацией 150 мМ, а во внутреннюю — 15 мМ. Натрий будет двигаться во внутреннюю ячейку, ривоноважний потенциал для него составит примерно 62 мВ.
Количество ионов, должна диффундировать для генерации электрического потенциала очень невелика (примерно 10 -12 моль К + на 1 см 2 мембраны), этот факт имеет два важных последствия. Во-прешь, это означает, что концентрации ионов, которые могут проникать через мембрану, остаются стабильными снаружи и внутри клетки, даже после того как их движение обеспечил утоврення электрического потенциала. Во-вторых, мизерные потоки ионов через мембрану, потирбно для установления потенциала, не нарушают электронейтральности цитоплазмы и внеклеточной жидкости в целом, распределение зарядов происходит только в области, непосредственно прилегающей к плазматической мембраны.
Уравнение Нернста
Равновесный потенциал для определенного иона, например для калия, можно рассчитать по уравнению Нернста, что выглядит так:
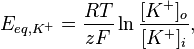 ,
,
где R — универсальная газовая постоянная, Т — абслоютна температура (по шкале Кельвина), z — заряд иона, F — число Фарадея, [K +] o, [K +] i — концентрация калия снаружи и внутри клетки соответственно. Поскольку описанные процессы происходят при температуре тела — 310 ° К, а десятичными логарифмами в исчислении пользоваться легче чем натуральными, это уравнение превращают следующим образом:
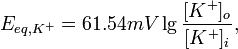
Подставляя концентрации К + в уравнение Нернста получаем равновесный потенциал для калия, составляет -90 мВ. Поскольку по нулевой потенциал принимается внешняя сторона мембраны, то знак минус означает, что в условиях равновесного калиевого потенциала внутренняя Сторн мембраны сравнительно более электроотрицательным. Аналогичные расчеты можно провести и для равновесного Натиева потенциала, он составляет 62 мВ.
Уравнения Голдмана
Хотя равновесный потенциал для ионов калия составляет -90 мВ, МПС нейрона несколько менее отрицательный. Эта разница отражает незначительное но постоянное следование ионов Na + через мембрану в состоянии покоя. Поскольку концентрационный градиент для натрия противоположный такого для калия, Na + движется внутрь клетки и сдвигает суммарный заряд на внутренней стороне мембраны в положительную сторону. На самом деле МПС нейрона составляет от -60 до -80 мВ. Это значение значительно ближе к Е K чем до Е Na, потому что в состоянии покоя в нейроне открыто много калиевых каналов и очень мало натриевых. Также на встанвлення МПС влияет движение ионов хлора. В 1943 году Дэвид Голдаман предложил усовершенствовать уравнение Нернста так, чтобы оно отражало влияние различных ионов на мембарнний потенциал, в этом уравнении учитывается относительная проницаемость мембраны для каждого типа ионов:
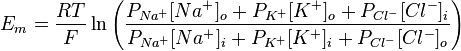
где R — универсальная газовая постоянная, Т — абслоютна температура (по шкале Кельвина), z — заряд иона, F — число Фарадея, [ион] o, [ион] i — концентрации ионов внутри и внутри клеток, Р — относительная проницаемость мембраны для соответствующего иона. Значение заряда в данном уравнении не сохраняется, но оно учтено — для хлора внешняя и внутренняя концентрация поменяны местами, так как его заряд 1.
Значение мембранного потенциала покоя для различных тканей
- Разделенными мышцы -95 мВ;
- Непосмугованих мышцы -50 мВ;
- Астроглией от -80 до -90 мВ;
- Нейроны -70 мВ.
Роль натрий-калиевого насоса в формировании МПС
Мембранный потенциал покоя может существовать только при условии неравномерного распределения ионов, обеспечивается функционированием натрий-калиевого насоса. Кроме того, этот белок делает также электрогенных властовости — он переносит 3 катионы натрия в обмен на 2 ионы калия, движущихся внутрь клетки. Таким образом, Na + -K + -АТФазы снижает МПС на 5-10 мВ. Подавление деятельности этого белка приводит к незначительному (на 5-10 мВ) мгновенного повышения мембранного потенциала, после чего он еще некоторое время будет существовать на достаточно стабильном уровне, пока будут сохраняться градиенты концентраций Na + и K +. Впоследствии эти градиенты начнут уменьшаться, вследствие проникнсоти мембраны к ионам, и через несколько десятков минут электрический потенциал на мембране исчезнет.
http://megalektsii.ru/s23176t4.html
http://info-farm.ru/alphabet_index/m/membrannyjj-potencial-pokoya.html