IV. Уравнение прямой, проходящей через данную точку в заданном направлении. Пучок прямых.
VI. Уравнение прямой, проходящей через две данные точки
VI.1.2. Распад высших жирных кислот
VI.2.2. Биосинтез триглицеридов
VIII.3. Биосинтез фосфатидов
XXIV. Нормы естественной убыли сыров жирных, полужирных, и низкожирных, упакованных в паро- и влагонепроницаемые полимерные пленки, при хранении в камерах холодильников
А) Мономолекулярлы реакция
Агглютинация реакциясы
Адиабатический процесс. Уравнение Пуассона.
Аналогично протекает реакция HgCl2 с сульфидом натрия Na2S.
Кроме пищевых жиров источником жирных кислот в организме служит их синтез из глюкозы. Непосредственным предшественником жирных кислот при их синтезе в организме является ацетил-КоА, т. е. то же вещество, которое образуется при р-окислении жирных кислот. Несмотря на то, что все реакции р-окисления обратимы, они не используются для синтеза жирных кислот.
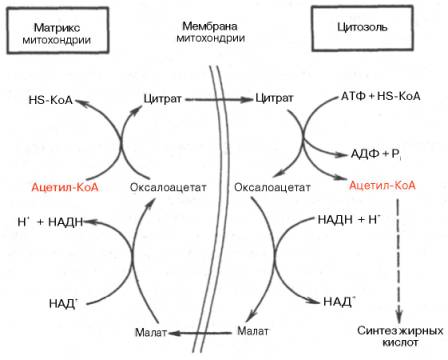
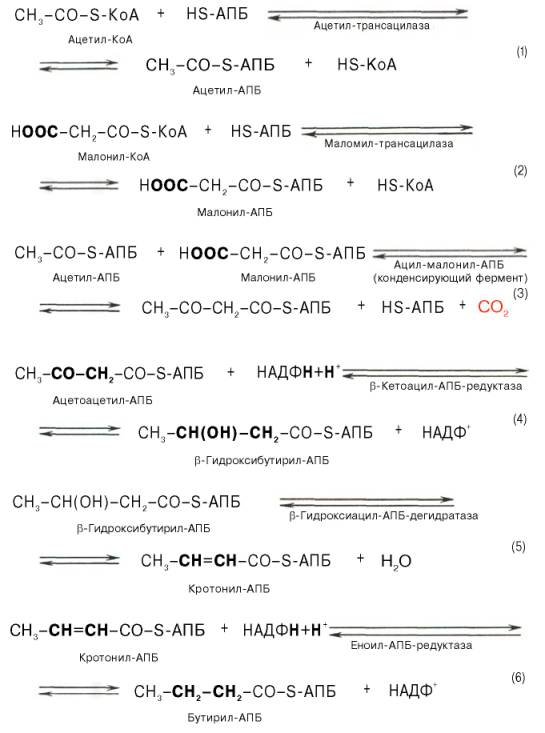
Ацетил-КоА для синтеза жирных кислот образуется путем окислительного де-карбоксилирования пирувата. Кроме того, окисление и синтез жирных кислот разделены в пространстве: окисление происходит в митохондриях, а синтез — в цитозоле.Перенос ацетильных остатков из митохондрий в цитозоль. Пируватдегидрогеназный комплекс локализован на внутренней поверхности внутренней мембраны митохондрий, и ацетил-КоА освобождается в матрикс митохондрий. Для синтеза жирных кислот ацетил-КоА должен быть перенесен в цитозоль. Мембрана митохондрий непроницаема для ацетил-КоА, и перенос ацетильного остатка в цитозоль происходит при участии челночного механизма (рис. 10.5). Пируват, образующийся из глюкозы в цитозоле, поступает в митохондрии, где частью превращается в ацетил-КоА (окислительное декарбоксилирование), частью — в оксалоацетат (при действии пи-руваткарбоксилазы). Затем из этих веществ образуется цитрат, для которого, как и для пирувата, есть белок-переносчик в мембране митохондрий. Образование малонил-КоА. Преобладающая часть ацетил-КоА, используемого для синтеза жирных кислот, вначале превращается в малонил-КоА при действии ацетил-КоА-карбоксилазы. Синтез пальмитиновой кислоты. Пальмитилсинтаза обладает каталитической активностью, в результате которой ацетильный и малонильный остатки переносятся на SH-группу пантотеновой кислоты (ацилтрансферазная активность) (рис. 10.7, реакции 1 и 2). Далее в реакции 3 ацетильный остаток переносится на место карбоксильной группы малонильного остатка; карбоксильная группа при этом отщепляется в виде С02 (реакция конденсации двух ацетильных остатков). Затем последовательно происходят восстановление р-карбонильной группы (реакция 4), отщепление воды с образованием двойной связи между а- и р-углеродны-ми атомами (реакция 5), восстановление (гидрирование) двойной связи (реакция 6). В результате получается остаток четырехуглеродной жирной кислоты, соединенный с ферментом (бутирил-Е). Все эти реакции катализируются разными активными центрами одного белка: как мы уже отмечали, пальмитилсинтаза — многофункциональный фермент. Субъединица пальмитилсинтазы представляет собой доменный белок, каждый домен которого катализирует одну из шести указанных реакций. Промежуточные продукты остаются постоянно связанными с ферментом через пантотеновую кислоту, перемещаясь на этой «привязи» из одного активного центра в другой.
Дата добавления: 2015-04-11 ; просмотров: 37 ; Нарушение авторских прав
БИОСИНТЕЗ НАСЫЩЕННЫХ ЖИРНЫХ КИСЛОТ
В настоящее время в достаточной степени изучен механизм биосинтеза жирных кислот в организме животных и человека, а также катализирующие этот процесс ферментные системы. Синтез жирных кислот протекает в цитоплазме клетки. В митохондриях в основном происходит удлинение существующих цепей жирных кислот. Установлено, что в цитоплазме печеночных клеток синтезируется пальмитиновая кислота (16 углеродных атомов), а в митохондриях этих клеток из уже синтезированной в цитоплазме клетки пальмитиновой кислоты или из жирных кислот экзогенного происхождения, т.е. поступающих из кишечника, образуются жирные кислоты, содержащие 18, 20 и 22 углеродных атома.
Иными словами, митохондриальная система биосинтеза жирных кислот, включающая несколько модифицированную последовательность реакций β-окисления, осуществляет только удлинение существующих в организме среднецепочечных жирных кислот, в то время как полный биосинтез пальмитиновой кислоты из ацетил-КоА активно протекает в цитозоле, т.е. вне митохондрий, по совершенно другому пути.
Внемитохондриальная система биосинтеза de novo жирных кислот (ли-погенез). Эта система находится в растворимой (цитозольной) фракции клеток многих органов, в частности печени, почек, мозга, легких, молочной железы, а также в жировой ткани. Биосинтез жирных кислот протекает с участием НАДФН, АТФ, Мn 2+ и НСО3 – (в качестве источника СО2); субстратом является ацетил-КоА, конечным продуктом – пальмитиновая кислота. Потребности в кофакторах процессов биосинтеза и β-окисления жирных кислот значительно различаются.
Как отмечалось, строительным блоком для синтеза жирных кислот в цитозоле клетки служит ацетил-КоА, который в основном поступает из митохондрий. Было выявлено, что цитрат стимулирует синтез жирных кислот в цитозоле клетки. Известно также, что образующийся в митохондриях в процессе окислительного декарбоксилирования пирувата и окисления жирных кислот ацетил-КоА не может диффундировать в цитозоль клетки, так как митохондриальная мембрана непроницаема для данного субстрата. Поэтому вначале внутримитохондриальный ацетил-КоА взаимодействует с оксалоацетатом, в результате чего образуется цитрат. Реакция катализируется ферментом цитрат-синтазой. Образовавшийся цитрат переносится через мембрану митохондрий в цитозоль при помощи специальной трикарбоксилаттранспортирующей системы.
В цитозоле цитрат реагирует с HS-KoA и АТФ, вновь распадаясь на ацетил-КоА и оксалоацетат. Эта реакция катализируется АТФ-цитратлиазой. Уже в цитозоле оксалоацетат при участии цитозольной малатдегидрогеназы восстанавливается до малата. Последний при помощи дикарбокси-латтранспортирующей системы возвращается в митохондриальный матрикс, где окисляется до оксалоацетата, завершая тем самым так называемый челночный цикл:
Существует еще один путь переноса внутримитохондриального аце-тил-КоА в цитозоль клетки – с участием карнитина. Как отмечалось, кар-нитин играет роль переносчика ацильных групп из цитозоля в митохондрии при окислении жирных кислот. По-видимому, он может выполнять эту роль и в обратном процессе, т.е. в переносе ацильных радикалов, в том числе ацетильного радикала, из митохондрий в цитозоль клетки. Однако, когда речь идет о синтезе жирных кислот, данный путь переноса ацетил-КоА не является главным.
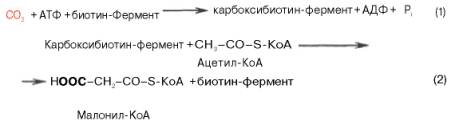
Образование малонил-КоА. Первой реакцией биосинтеза жирных кислот является карбоксилирование ацетил-КоА, для чего требуются бикарбонат, АТФ, ионы марганца. Катализирует эту реакцию фермент ацетил-КоА-карбоксилаза. Фермент содержит в качестве простетической группы биотин. Авидин – ингибитор биотина угнетает эту реакцию, как и синтез жирных кислот в целом.
Установлено, что ацетил-КоА-карбоксилаза состоит из переменного числа одинаковых субъединиц, каждая из которых содержит биотин, биотинкарбоксилазу, карбоксибиотинпереносящий белок, транскарбоксилазу, а также регуляторный ал-лостерический центр, т.е. представляет собой полиферментный комплекс.
Реакция протекает в два этапа: I – карбоксилирование биотина с участием АТФ и II – перенос карбоксильной группы на ацетил-КоА, в результате чего образуется малонил-КоА:
Малонил-КоА представляет собой первый специфический продукт биосинтеза жирных кислот. В присутствии соответствующей ферментной системы малонил-КоА быстро превращается в жирные кислоты.
Энзиматические системы, осуществляющие синтез жирных кислот, называются жирно-кислотными синтетазами. Они широко встречаются в природе и могут быть изолированы из различных одноклеточных организмов, растений и животных тканей.
Жирно-кислотные синтетазы делятся на 2 группы. К первой группе относятся полиэнзимные, не поддающиеся фракционированию комплексы с мол. м. порядка 500000, в которых все индивидуальные энзимы собраны в компактную структуру. В частности, в эту группу входят жирно-кислотные синтетазы животных тканей и дрожжей.
Вторая группа включает жирно-кислотные синтетазы, из которых отдельные энзимы могут быть выделены методами белкового фракционирования. Такие синтетазы встречаются у ряда микроорганизмов (в частности, у E.coli) и растений. Иными словами, в этих случаях все индивидуальные ферменты синтетазной системы находятся в виде автономных полипептидов.
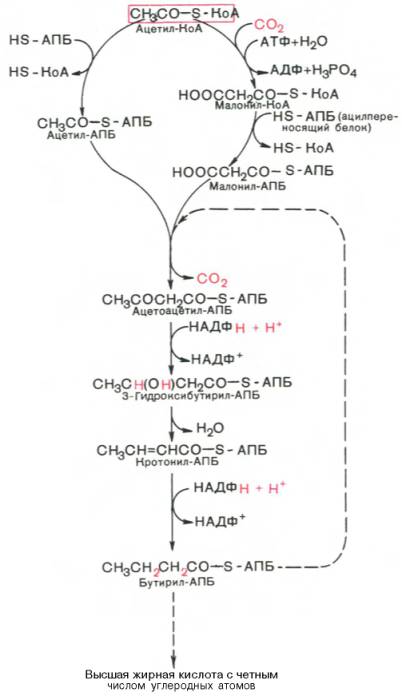
Мультиферментный комплекс, называемый синтетазой (синтазой) жирных кислот, состоит из 6 ферментов, связанных с так называемым ацилпереносящим белком (АПБ). Этот белок относительно термостабилен, имеет две свободные HS-группы (цистеина и фосфопантетеинового остатка, присоединенного к ОН-группе серина) и вовлекается в процесс синтеза высших жирных кислот практически на всех его этапах. Мол. масса АПБ составляет около 10000. Данный белок в синтетазной системе выполняет роль КоА. Заметим, что в животных тканях не удалось обнаружить свободного АПБ, подобного микробному. Из печени выделен полиэнзимный комплекс, содержащий все энзимы, необходимые для синтеза жирных кислот. Энзимы комплекса настолько прочно связаны друг с другом, что все попытки изолировать их в индивидуальном виде не увенчались успехом. Приводим последовательность реакций, происходящих при синтезе жирных кислот:
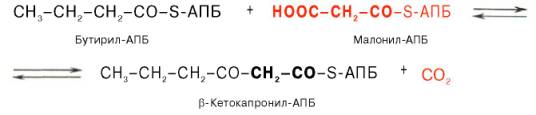
Далее цикл реакций повторяется. Допустим, что идет синтез пальмитиновой кислоты (С16). В этом случае образованием бутирил-АПБ завершается лишь первый из 7 циклов, в каждом из которых началом является присоединение молекулы малонил-АПБ к карбоксильному концу растущей цепи жирной кислоты. При этом отщепляется дистальная карбоксильная группа малонил-АПБ в виде СО2. Например, образовавшийся в первом цикле бутирил-АПБ взаимодействует с малонил-АПБ:
Завершается синтез жирной кислоты отщеплением HS-АПБ от ацил-АПБ под влиянием фермента деацилазы. Например:
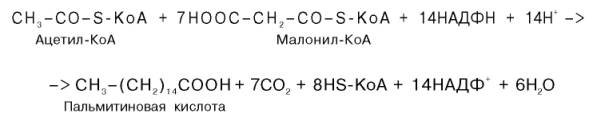
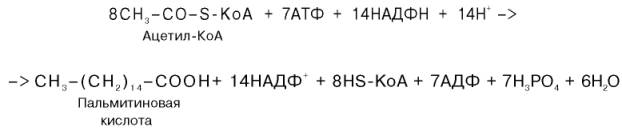
Суммарное уравнение синтеза пальмитиновой кислоты можно записать так:
Или, учитывая, что на образование одной молекулы малонил-КоА из ацетил-КоА расходуются одна молекула АТФ и одна молекула СО2, которая затем отщепляется, суммарное уравнение можно представить в следующем виде:
Основные этапы биосинтеза жирных кислот можно представить в виде схемы:
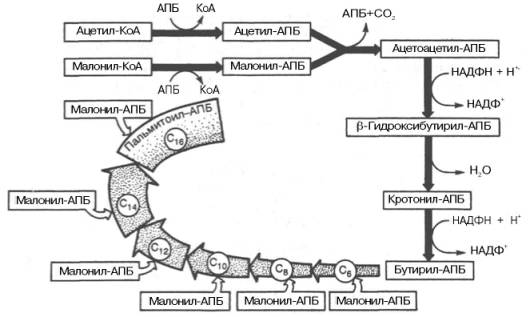
В общем виде синтез жирных кислот у кишечной палочки представлен на рис. 11.4. Последовательность и характер реакций в синтезе жирных кислот, начиная с образования β-кетоацил-АПБ (на рис. 11.4 – ацетоацетил-АПБ) и кончая завершением одного цикла удлинения цепи на два углеродных атома, являются как бы обратными реакциями окисления жирных кислот. На самом деле пути синтеза и окисления жирных кислот не пересекаются даже частично. Это становится очевидным, если принять во внимание некоторые особенности синтеза и окисления жирных кислот.
Рис. 11.4. Синтез пальмитиновой кислоты у кишечной палочки при участии одной молекулы ацетил-КоА и 7 молекул малонил-КоА. Подробно представлен первый цикл синтеза — образование бутирил-АПБ. Остальные 6 циклов аналогичны первому.
По сравнению с β-окислением биосинтез жирных кислот имеет ряд характерных особенностей: синтез жирных кислот в основном осуществляется в цитозоле клетки, а окисление – в митохондриях; участие в процессе биосинтеза жирных кислот малонил-КоА, который образуется путем связывания СО2 (в присутствии биотин-фермента и АТФ) с ацетил-КоА; на всех этапах синтеза жирных кислот принимает участие ацилпереносящий белок (HS-АПБ); при биосинтезе образуется D(–)-изомер 3-гидроксикис-лоты, а не L(+)-изомер, как это имеет место при β-окислении жирных кислот; необходимость для синтеза жирных кислот кофермента НАДФН. Последний в организме частично (на 50%) образуется в реакциях пен-тозофосфатного цикла, частично – в других реакциях, в частности в реакциях:
Образование ненасыщенных жирных кислот. Элонгация жирных кислот. В отличие от растительных тканей ткани животных обладают весьма ограниченной способностью превращать насыщенные жирные кислоты в ненасыщенные.
Установлено, что две наиболее распространенные мононенасыщенные жирные кислоты – пальмитоолеиновая и олеиновая – синтезируются из пальмитиновой и стеариновой кислот.
Эти превращения протекают в микросомах клеток печени и жировой ткани при участии молекулярного кислорода, восстановленной системы пиридиновых нуклеотидов и цитохрома b5. Превращению подвергаются только активированные формы пальмитиновой и стеариновой кислот. Ферменты, участвующие в этих превращениях, получили название деса-тураз.
Наряду с десатурацией жирных кислот (образование двойных связей) в микросомах происходит и их удлинение (элонгация), причем оба эти процесса могут сочетаться и повторяться. Удлинение цепи жирной кислоты происходит путем последовательного присоединения к соответствующему ацил-КоА двууглеродных фрагментов при участии малонил-КоА и НАДФН. Энзиматическая система, катализирующая удлинение жирных кислот, получила название элонгазы. На схеме представлены пути превращения пальмитиновой кислоты в реакциях десатурации и элонгации.
Acetyl
Наведите курсор на ячейку элемента, чтобы получить его краткое описание.
Чтобы получить подробное описание элемента, кликните по его названию.
H +
Li +
K +
Na +
NH4 +
Ba 2+
Ca 2+
Mg 2+
Sr 2+
Al 3+
Cr 3+
Fe 2+
Fe 3+
Ni 2+
Co 2+
Mn 2+
Zn 2+
Ag +
Hg 2+
Pb 2+
Sn 2+
Cu 2+
OH —
Р
Р
Р
Р
Р
М
Н
М
Н
Н
Н
Н
Н
Н
Н
Н
—
—
Н
Н
Н
F —
Р
М
Р
Р
Р
М
Н
Н
М
М
Н
Н
Н
Р
Р
Р
Р
Р
—
Н
Р
Р
Cl —
Р
Р
Р
Р
Р
Р
Р
Р
Р
Р
Р
Р
Р
Р
Р
Р
Р
Н
Р
М
Р
Р
Br —
Р
Р
Р
Р
Р
Р
Р
Р
Р
Р
Р
Р
Р
Р
Р
Р
Р
Н
М
М
Р
Р
I —
Р
Р
Р
Р
Р
Р
Р
Р
Р
Р
?
Р
?
Р
Р
Р
Р
Н
Н
Н
М
?
S 2-
М
Р
Р
Р
Р
—
—
—
Н
—
—
Н
—
Н
Н
Н
Н
Н
Н
Н
Н
Н
HS —
Р
Р
Р
Р
Р
Р
Р
Р
Р
?
?
?
?
?
Н
?
?
?
?
?
?
?
SO3 2-
Р
Р
Р
Р
Р
Н
Н
М
Н
?
—
Н
?
Н
Н
?
М
М
—
Н
?
?
HSO3 —
Р
?
Р
Р
Р
Р
Р
Р
Р
?
?
?
?
?
?
?
?
?
?
?
?
?
SO4 2-
Р
Р
Р
Р
Р
Н
М
Р
Н
Р
Р
Р
Р
Р
Р
Р
Р
М
—
Н
Р
Р
HSO4 —
Р
Р
Р
Р
Р
Р
Р
Р
—
?
?
?
?
?
?
?
?
?
?
Н
?
?
NO3 —
Р
Р
Р
Р
Р
Р
Р
Р
Р
Р
Р
Р
Р
Р
Р
Р
Р
Р
Р
Р
—
Р
NO2 —
Р
Р
Р
Р
Р
Р
Р
Р
Р
?
?
?
?
Р
М
?
?
М
?
?
?
?
PO4 3-
Р
Н
Р
Р
—
Н
Н
Н
Н
Н
Н
Н
Н
Н
Н
Н
Н
Н
Н
Н
Н
Н
CO3 2-
Р
Р
Р
Р
Р
Н
Н
Н
Н
?
?
Н
?
Н
Н
Н
Н
Н
?
Н
?
Н
CH3COO —
Р
Р
Р
Р
Р
Р
Р
Р
Р
—
Р
Р
—
Р
Р
Р
Р
Р
Р
Р
—
Р
SiO3 2-
Н
Н
Р
Р
?
Н
Н
Н
Н
?
?
Н
?
?
?
Н
Н
?
?
Н
?
?
Растворимые (>1%)
Нерастворимые (
Спасибо! Ваша заявка отправлена, преподаватель свяжется с вами в ближайшее время.
Вы можете также связаться с преподавателем напрямую:
8(906)72 3-11-5 2
Скопируйте эту ссылку, чтобы разместить результат запроса » » на другом сайте.
Изображение вещества/реакции можно сохранить или скопировать, кликнув по нему правой кнопкой мыши.
Если вы считаете, что результат запроса » » содержит ошибку, нажмите на кнопку «Отправить».
Этим вы поможете сделать сайт лучше.
К сожалению, регистрация на сайте пока недоступна.
На сайте есть сноски двух типов:
Подсказки — помогают вспомнить определения терминов или поясняют информацию, которая может быть сложна для начинающего.
Дополнительная информация — такие сноски содержат примечания или уточнения, выходящие за рамки базовой школьной химии, нужны для углубленного изучения.
Здесь вы можете выбрать параметры отображения органических соединений.
 Ацетил-КоА для синтеза жирных кислот образуется путем окислительного де-карбоксилирования пирувата. Кроме того, окисление и синтез жирных кислот разделены в пространстве: окисление происходит в митохондриях, а синтез — в цитозоле.Перенос ацетильных остатков из митохондрий в цитозоль. Пируватдегидрогеназный комплекс локализован на внутренней поверхности внутренней мембраны митохондрий, и ацетил-КоА освобождается в матрикс митохондрий. Для синтеза жирных кислот ацетил-КоА должен быть перенесен в цитозоль. Мембрана митохондрий непроницаема для ацетил-КоА, и перенос ацетильного остатка в цитозоль происходит при участии челночного механизма (рис. 10.5). Пируват, образующийся из глюкозы в цитозоле, поступает в митохондрии, где частью превращается в ацетил-КоА (окислительное декарбоксилирование), частью — в оксалоацетат (при действии пи-руваткарбоксилазы). Затем из этих веществ образуется цитрат, для которого, как и для пирувата, есть белок-переносчик в мембране митохондрий. Образование малонил-КоА. Преобладающая часть ацетил-КоА, используемого для синтеза жирных кислот, вначале превращается в малонил-КоА при действии ацетил-КоА-карбоксилазы. Синтез пальмитиновой кислоты. Пальмитилсинтаза обладает каталитической активностью, в результате которой ацетильный и малонильный остатки переносятся на SH-группу пантотеновой кислоты (ацилтрансферазная активность) (рис. 10.7, реакции 1 и 2). Далее в реакции 3 ацетильный остаток переносится на место карбоксильной группы малонильного остатка; карбоксильная группа при этом отщепляется в виде С02 (реакция конденсации двух ацетильных остатков). Затем последовательно происходят восстановление р-карбонильной группы (реакция 4), отщепление воды с образованием двойной связи между а- и р-углеродны-ми атомами (реакция 5), восстановление (гидрирование) двойной связи (реакция 6). В результате получается остаток четырехуглеродной жирной кислоты, соединенный с ферментом (бутирил-Е). Все эти реакции катализируются разными активными центрами одного белка: как мы уже отмечали, пальмитилсинтаза — многофункциональный фермент. Субъединица пальмитилсинтазы представляет собой доменный белок, каждый домен которого катализирует одну из шести указанных реакций. Промежуточные продукты остаются постоянно связанными с ферментом через пантотеновую кислоту, перемещаясь на этой «привязи» из одного активного центра в другой.
Ацетил-КоА для синтеза жирных кислот образуется путем окислительного де-карбоксилирования пирувата. Кроме того, окисление и синтез жирных кислот разделены в пространстве: окисление происходит в митохондриях, а синтез — в цитозоле.Перенос ацетильных остатков из митохондрий в цитозоль. Пируватдегидрогеназный комплекс локализован на внутренней поверхности внутренней мембраны митохондрий, и ацетил-КоА освобождается в матрикс митохондрий. Для синтеза жирных кислот ацетил-КоА должен быть перенесен в цитозоль. Мембрана митохондрий непроницаема для ацетил-КоА, и перенос ацетильного остатка в цитозоль происходит при участии челночного механизма (рис. 10.5). Пируват, образующийся из глюкозы в цитозоле, поступает в митохондрии, где частью превращается в ацетил-КоА (окислительное декарбоксилирование), частью — в оксалоацетат (при действии пи-руваткарбоксилазы). Затем из этих веществ образуется цитрат, для которого, как и для пирувата, есть белок-переносчик в мембране митохондрий. Образование малонил-КоА. Преобладающая часть ацетил-КоА, используемого для синтеза жирных кислот, вначале превращается в малонил-КоА при действии ацетил-КоА-карбоксилазы. Синтез пальмитиновой кислоты. Пальмитилсинтаза обладает каталитической активностью, в результате которой ацетильный и малонильный остатки переносятся на SH-группу пантотеновой кислоты (ацилтрансферазная активность) (рис. 10.7, реакции 1 и 2). Далее в реакции 3 ацетильный остаток переносится на место карбоксильной группы малонильного остатка; карбоксильная группа при этом отщепляется в виде С02 (реакция конденсации двух ацетильных остатков). Затем последовательно происходят восстановление р-карбонильной группы (реакция 4), отщепление воды с образованием двойной связи между а- и р-углеродны-ми атомами (реакция 5), восстановление (гидрирование) двойной связи (реакция 6). В результате получается остаток четырехуглеродной жирной кислоты, соединенный с ферментом (бутирил-Е). Все эти реакции катализируются разными активными центрами одного белка: как мы уже отмечали, пальмитилсинтаза — многофункциональный фермент. Субъединица пальмитилсинтазы представляет собой доменный белок, каждый домен которого катализирует одну из шести указанных реакций. Промежуточные продукты остаются постоянно связанными с ферментом через пантотеновую кислоту, перемещаясь на этой «привязи» из одного активного центра в другой.