Рост числа клеток популяции описывается уравнением
Теория : Нелимитированный рост численности популяции при постоянных условиях среды описывается дифференциальным уравнением Мальтуса (Ризниченко, Рубин, 1993) :
,(1.1)
где x – численность (плотность) популяции, t – время, r – мальтузианский параметр, имеющий смысл удельной скорости роста. Размерность мальтузианского параметра – [время -1 ]. Решением данного уравнения является выражение:
,(1.2)
где x 0 – начальная численность популяции.
Условия : С целью изучения динамики эвтрификации водоемов, загрязненных минеральными удобрениями, в пяти прудах моделировали размножение синезеленых водорослей в нелимитированных условиях. Полученые данные об изменении численности популяции водорослей в каждом пруду представлены в таблице 1.1.
Таблица 1.1
| Время, час t | Титр клеток водорослей, кл./мл | ||||
| Пруд 1 x1 | Пруд 2 x2 | Пруд 3 x3 | Пруд 4 x4 | Пруд 5 x5 | |
| 0 | 135 | 171 | 60 | 252 | 106 |
| 24 | 245 | 270 | 113 | 371 | 201 |
| 48 | 374 | 491 | 186 | 710 | 275 |
| 72 | 545 | 693 | 269 | 1088 | 451 |
| 96 | 839 | 1163 | 447 | 1772 | 689 |
| 120 | 1544 | 1788 | 796 | 2534 | 1304 |
| 144 | 2392 | 3460 | 1024 | 4842 | 2161 |
| 168 | 3433 | 4704 | 2131 | 6478 | 3386 |
| 192 | 6586 | 8526 | 3107 | 10429 | 5326 |
| 216 | 10129 | 13198 | 4351 | 19953 | 8928 |
Вопросы: На основе этих данных определить для популяции водорослей в каждом пруду значение мальтузианского параметра r (удельной скорости размножения) и период удвоения T . Найти также соответствующие медианы по полученным выборкам r и T .
Решение: Нелимитированный рост численности популяции описывается экспоненциальной функцией Мальтуса:
где x 0 – начальная численность популяции, r – мальтузианский параметр, t – время (см. рис.1.1).
Рис.1.1. Экспоненциальная зависимость численности популяции от времени при условии нелимитированного роста
Для обработки экспериментальных данных используем логарифмическую форму этого уравнения:
(1.4)
С этой целью каждый элемент 2-6 столбцов необходимо разделить на соответствующий начальный элемент и взять натуральный логарифм этого отношения (см. таблицу 1.2).
Далее, для каждого пруда на основе таблицы 1.2 следует построить график, где по оси абсциcc откладывается время, а по оси ординат – соответствующее значение ln (x t /x 0 ), как это показано на рис.1.2. Экспериментальные данные должны сгруппироваться около прямой линии, тангенс угла которой и есть искомое значение мальтузианского параметра r .
Таблица 1.2
| Время, час | ln ( x 1 t /x 1 0 ) | ln ( x 2 t /x 2 0 ) | ln ( x 3 t /x 3 0 ) | ln ( x 4 t /x 4 0 ) | ln ( x 5 t /x 5 0 ) |
| 0,00 | 0,00 | 0,00 | 0,00 | 0,00 | 0,00 |
| 24,00 | 0,60 | 0,46 | 0,63 | 0,39 | 0,64 |
| 48,00 | 1,02 | 1,06 | 1,13 | 1,04 | 0,95 |
| 72,00 | 1,40 | 1,40 | 1,50 | 1,46 | 1,45 |
| 96,00 | 1,83 | 1,92 | 2,01 | 1,95 | 1,87 |
| 120,00 | 2,44 | 2,35 | 2,59 | 2,31 | 2,51 |
| 144,00 | 2,87 | 3,01 | 2,84 | 2,96 | 3,01 |
| 168,00 | 3,24 | 3,31 | 3,57 | 3,25 | 3,46 |
| 192,00 | 3,89 | 3,91 | 3,95 | 3,72 | 3,92 |
| 216,00 | 4,32 | 4,35 | 4,28 | 4,37 | 4,43 |
Рис.1.2. Линеаризация экспоненциальной зависимости, представленной на рис.1.1, и определение мальтузианского параметра r .
Значение периода удвоения T находим по формуле:
(1.5)
Результаты обработки данных в этой задаче представлены в таблице 1.3.
Таблица 1.3
| Пруд 1 | Пруд 2 | Пруд 3 | Пруд 4 | Пруд 5 | |
| r , час -1 | 0.0197 | 0.0201 | 0.0198 | 0.0198 | 0.0204 |
| 35.19 | 34.48 | 35 | 35 | 33.98 |
Медиану определяем обычным способом как срединное значение в выборке:
Ме( r )= 0.0198 час -1 , Ме( T )= 35 час.
Тема 2. Плотностно-зависимый рост популяций. Логистическое уравнение Ферхюльста.
Теория : При ограничении процессов размножения организмов в популяции каким либо ресурсом, например количеством доступной пищи, удельная скорость роста популяции зависит от ее численности (плотности). Математические модели, учитывающие данный эффект, называются моделями плотностно-зависимого роста. Логистическое уравнение Ферхюльста является простейшей моделью из этого ряда. В данной модели предполагается, что удельная скорость роста популяции линейно уменьшается с ростом численности, и имеется также некая предельная численность популяции, K , при достижении которой добавление к популяции новых особей возможно лишь при условии определенной гибели уже имеющихся особей. Эта предельная численность K носит название емкость среды. Данный параметр известен также как ресурсный. Уравнение Ферхюльста имеет следующий вид:
Здесь r – мальтузианский параметр, K – ресурсный параметр. Уравнение Ферхюльста имеет аналитическое решение:
где x 0 – начальная численность популяции. Пример логистической популяционной динамики приведен на рис. 2.1
Рис.2.1. Логистический рост популяции.
r = 0.07 год -1 , K = 1000 особей, >x 0 = 10 особей.
Условия: В 1937 г. на остров Протекшен у побережья штата Вашингтон завезли двух самцов и шесть самок обыкновенного фазана ( Phasianus colchicus ). За пять лет численность популяции фазана достигла 1325 размножающихся особей, т.е. возросла в 166 раз. Судя по некоторым признакам, рост этой популяции происходит по логистической кривой. От двух самцов и шести самок, привезенных на остров в 1937 г., она возросла до 30 особей к весне 1938 г., до 81 – в 1939 г., до 282 – в 1940 г., 641 – в 1941 г., 1194 – в 1942 г. и до 1898 – в 1943 г., когда на остров прибыли военные части, и солдаты начали охотиться на фазанов (рис.2.2). Однако, создается впечатление, что к 1943 г. численность популяции фазанов начитает стабилизироваться (Солбриг,Солбриг, 1982).
Рис.2.2. Измнение численности популяции фазанов на острове Протекшен.
Вопросы:
- Показать, что в данной популяции фазанов наблюдается плотностно-зависимый рост численности.
- Используя приведенные данные о росте численности популяции фазанов найти для этой популяции значения параметров логистическго уравнения Ферхюльста: мальтузианского параметра r и ресурсного параметра К .
Решение:
1. Для ответа на первый вопрос представим данные в координатах < ln ( x ) , t >, где x – численность популяции, t – время. При плотностно-зависимом росте данные в указанных координатах должны отклонятся от прямой линии с течением времени, что и наблюдается в нашем случае (рис.2.3). Это свидетельствует о том, что удельная скорость роста уменьшается с увеличением численности.
2. Решение второго вопроса разобьем на два этапа. На первом этапе используем приближенное выражение, получаемое из уравнения Ферхюльста (2.1):
Рис.2.3. Изменение численности популяции фазанов в полулогарифмических координатах
Для этого на основе первичных данных рассчитаем дополнительные переменные и , как это указано в таблице 2.1, и представим их в виде графика, где по оси абсцисс будем откладывать , а по оси ординат — (рис.2.4).
ln ( x )
Рис.2.4. Определение параметра K по дополнительным переменным, представленным в таб. 2.1.
Точки на графике группируются около прямой линии, которая согласно выражению (2.3) пересекает ось абсцисс в точке, соответствующей величине K (рис.2.4). В рассматриваемом случае величина параметра K равна 2500.
Рис. 2.5. Определение параметра r по данным из таблицы 2.1.
Для нахождения параметра r воспользуемся интегральной формой уравнения Ферхюльста в логарифмическом виде:
На основе ряда x рассчитаем дополнительную переменную , как указано в таблице 2.1. и построим график для этой переменной против переменной t (рис. 2.5). Как видно, экспериментальные точки группируются около прямой линии, тангенс угла которой согласно выражению (2.4) дает значение параметра r . Для рассматриваемого случая tg a = 1.13. Учитывая размерность шкал получаем искомый ответ: r = 1.13 год -1 .
Условия: В Кузнецком Алатау на одном из охотучастков велась добыча соболя. Через некоторое время охотоведы заметили резкое снижение численности популяции. Когда численность соболя на участке упала ниже известной критической величины, был сделан вывод о неконтролируемом браконьерском промысле. Контролируемый промысел был остановлен, и был организован ежегодный учет численности популяции соболя. Данные представлены в таблице 2.2 и на рис.2.6.
| Время, год | Численность | Время, год | Численность |
| 0 | 1120 | 8 | 1940 |
| 1 | 1147 | 9 | 2072 |
| 2 | 1189 | 10 | 2173 |
| 3 | 1253 | 11 | 2245 |
| 4 | 1346 | 12 | 2293 |
| 5 | 1470 | 13 | 2324 |
| 6 | 1620 | 14 | 2343 |
| 7 | 1784 | 15 | 2355 |
Рис.2.6. Изменение численности популяции соболя после отмены контролируемого промысла
Вопросы: По данным, представленным в таблице 2.2. определите, сколько соболей в год браконьеры добывали на данном участке N p , каковы кинетические параметры популяции соболей на данном участке ( r и K ), какова критическая численность популяции в условиях браконьерства x кр. и какова критическая величина промысла N кр .
Решение: Будем описывать динамику этой популяции следующим уравнением (Базыкин, 1985):
Здесь r — мальтузианский параметр, K – ресурсный параметр, N p – браконьерский промысел.
Для решения на первом этапе используем приближенную форму данного уравнения:
С этой целью на основе данных о x i и t i рассчитаем дополнительные переменные и (см. таб.2.3.), а затем построим график в этих координатах (рис.2.7). Точки на графике должны лечь на кривую, перевернутую параболу, которая пересекает ось абсцисс в точках X 1 и X 2 , соответствующих стационарным решениям уравнения (2.5).
Для исследуемой популяции по графику (рис.2.7) определяем, что X 1 = 1086 и X 2 = 2373. Из этих данных далее определяем значение параметра K , т.к. из теории (Базыкин, 1985) известно, что K = X 1 + X 2 . В нашем случае K =1086 + 2373 = 3459 особей.
Знание параметра K и значений X 1 и X 2 позволяет нам использовать для дальнейшего решения задачи интегральную форму уравнения (2.5):
| Время, год t | Численность x | |||
| 0 | 1120 | -3,60 | ||
| 1 | 1146 | 1133 | 26,9 | -3,00 |
| 2 | 1189 | 1168 | 42,2 | -2,43 |
| 3 | 1253 | 1221 | 64,1 | -1,90 |
| 4 | 1345 | 1299 | 92,4 | -1,37 |
| 5 | 1469 | 1407 | 123 | -0,85 |
| 6 | 1620 | 1544 | 150 | -0,34 |
| 7 | 1783 | 1701 | 163 | 0,16 |
| 8 | 1939 | 1861 | 156 | 0,67 |
| 9 | 2072 | 2005 | 132 | 1,18 |
| 10 | 2173 | 2122 | 101 | 1,69 |
| 11 | 2245 | 2209 | 71,6 | 2,20 |
| 12 | 2292 | 2268 | 47,8 | 2,71 |
| 13 | 2323 | 2308 | 30,7 | 3,22 |
| 14 | 2342 | 2333 | 19,2 | 3,72 |
| 15 | 2354 | 2348 | 11,8 | 4,23 |
Рис.2.7. Определение параметров X 1 и X 2 , по данным из таблицы 2.3.
Рассчитаем соответствующую дополнительную переменную , как указано в таб.2.3. и построим график для этой переменной против переменной t (рис.2.8).
Рис.2.8. Определение параметра r по данным из таблицы 2.3.
Для этого графика tg a = 0.517. Далее определяем значение параметра r :
Учитывая размерность осей получаем r = 1.39 год -1 .
Значение параметра N p (интенсивность браконьерского промысла) находим из выражения: ,
Критическая численность популяции в условиях браконьерского промысла, согласно теории, равна стационарному решению X 1 , поэтому x кр. = 1086 особей. Критическую величину промысла N кр находим из соотношения особей.
Тема 3. Влияние концентрации лимитирующего субстрата на скорость размножения. Уравнение Моно.
Теория : Из множества необходимых для жизнедеятельности пищевых компонентов, субстратов, потребляемых организмами исследуемой популяции, субстрат, находящийся в наименьшей доступности и лимитирующий ростовые процессы, определяет скорость размножения. Этот принцип известен как принцип минимума Либиха. Количественная зависимость удельной скорости размножения r от количества (концентрации) лимитирующего субстрата S, как правило, имеет насыщающий характер и обычно описывается уравнением Моно (Печуркин, 1978; Ризниченко, Рубин, 1993):
Уравнение Моно имеет два параметра: r max – максимально возможная скорость размножения при избытке субстрата (размерность – [время] -1 ) и K S – коэффициент насыщения (размерность – аналогичная размерности субстрата). Согласно теории коэффициент насыщения численно равен той концентрации субстрата, при которой наблюдается половинная от максимальной скорость размножения. Типичный вид зависимости Моно представлен на рис.3.1.
Рис.3.1. Зависимость удельной скорости размножения от концентрации лимитирующего субстрата в соответствии с уравнением Моно
Условия: С целью определения кинетики размножения популяции бактерий Escherichia coli (кишечная палочка) изучали рост и размножение клеток данных бактерий при различных концентрациях лимитирующего субстрата, глицерина. Клетки выращивали при оптимальных условиях и через каждые полчаса измеряли оптическую плотность культуры (показатель, отражающий численность популяции). Полученные в результате эксперимента данные представлены в таблице 3.1. и на рис.3.2.
Таблица 3.1.
| Время час | Оптическая плотность культуры при различной концентрации субстрата (мг/л), Ex | ln ( Ex t /Ex 0 ) при различной концентрации субстрата (мг/л) | ||||||||
| 50 | 100 | 200 | 400 | 800 | 50 | 100 | 200 | 400 | 800 | |
| 0,00 | 0,12 | 0,12 | 0,09 | 0,14 | 0,10 | -0,05 | -0,01 | 0,01 | -0,01 | 0,03 |
| 0,50 | 0,16 | 0,15 | 0,12 | 0,19 | 0,17 | 0,22 | 0,20 | 0,32 | 0,31 | 0,53 |
| 1,00 | 0,17 | 0,20 | 0,18 | 0,27 | 0,26 | 0,27 | 0,50 | 0,67 | 0,66 | 0,96 |
| 1,50 | 0,24 | 0,24 | 0,24 | 0,42 | 0,36 | 0,63 | 0,68 | 0,96 | 1,10 | 1,29 |
| 2,00 | 0,29 | 0,34 | 0,31 | 0,59 | 0,48 | 0,80 | 1,03 | 1,22 | 1,43 | 1,57 |
| 2,50 | 0,34 | 0,44 | 0,49 | 0,82 | 0,75 | 0,95 | 1,29 | 1,69 | 1,77 | 2,02 |
| 3,00 | 0,40 | 0,59 | 0,61 | 1,32 | 1,06 | 1,12 | 1,59 | 1,91 | 2,24 | 2,36 |
| 3,50 | 0,53 | 0,82 | 0,94 | 1,81 | 1,75 | 1,41 | 1,93 | 2,34 | 2,56 | 2,86 |
| 4,00 | 0,63 | 1,04 | 1,36 | 2,41 | 2,35 | 1,57 | 2,16 | 2,72 | 2,84 | 3,16 |
| 4,50 | 0,78 | 1,34 | 1,79 | 3,62 | 3,96 | 1,79 | 2,42 | 2,99 | 3,25 | 3,68 |
Рис.3.2. Изменение оптической плотности культуры клеток E.coli при различной концентрации лимитирующего субстрата (глицерина). Здесь S50 – концентрация субстрата 50 мг/л,
S100 – концентрация субстрата 100 мг/л и т.д.
Вопросы: Определить на основе представленных экспериментальных данных кинетические параметры размножения клеток кишечной палочки (параметры уравнения Моно, r max и K S )
Решение: На первом этапе обработки экспериментальных данных определим удельную скорость роста r клеток кишечной палочки для каждой концентрации лимитирующего субстрата по алгоритму, рассмотренному в задаче 1.1. В качестве показателя, отражающего относительное изменение численности популяции во времени, будем использовать отношение соответствующих значений оптической плотности Ex t /Ex 0 (Перт,1978). Рассчитаем для каждой концентрации дополнительную переменную ln( Ex t /Ex 0 ) и построим график зависимости этой переменной от времени (таб.3.1. и рис.3.3). Экспериментальные точки для каждой концентрации должны сгруппироваться около соответствующей прямой линии, тангенс угла которой равен оценке r для этой концентрации.
Рис.3.3. Определение значений удельной скорости размножения r клеток E.coli для каждой концентрации лимитирующего субстрата. Обозначения те же, что и на рис.3.2.
Определенные таким образом значения r для каждой концентрации лимитирующего субстрата S представлены в таблице 3.2., а соответствующий график – на рис.3.4.
| Концентрация лимитирующего субстрата S , мг/л | 50 | 100 | 200 | 400 | 800 |
| Удельная скорость размножения r , час -1 | 0.405 | 0.55 | 0.67 | 0.73 | 0.78 |
Рис.3.4. Зависимость удельной скорости размножения исследуемой популяции кишечной палочки от концентрации лимитирущего субстрата, глицерина
Нетрудно видеть, что зависимость удельной скорости размножения от концентрации субстрата для исследуемой популяции имеет вид кривой с насыщением и по всей видимости описывается уравнением Моно (3.1).
Для нахождения параметров уравнения Моно воспользуемся методом Корниша-Боудена, “прямым линейным графиком”, разработанным специально для подобного рода зависимостей (Корниш-Боуден, 1979). С этой целью для каждой пары < S, r >из таб.3.2 на оси абсцисс влево будем откладывать значение – S , а на оси ординат – значение r . Далее через точки <- S, 0>и <0 , r >проводим прямую линию (рис.3.5), Таких линий на графике будет пять, по числу пар < S, r >. Данные прямые по теории Корниша-Боудена должны пересечься в одной точке, координаты которой дают искомые значения параметров уравнения Моно, K S и r max . В рассматриваемой задаче из-за наличия ошибок измерений точек пересечения прямых на графике (рис.3.5) больше, около десяти. В этом случае в качестве оценки рекомендуют брать медиану.
Рис.3.5. Определение кинетических параметров размножения клеток кишечной палочки K S и r max методом Корниша-Боудена на основе данных из таб.3.2.
Таким образом, по графику на рис.3.5. определяем, что для исследуемой популяции E.coli кинетические параметры размножения имеют следующие значения:
K S = 48 мг/л, r max = 0.83 час -1 .
Условия: Изучали кинетику размножения богомола. Исходя из предположения о пропорциональности между интенсивностью поедания пищи и скоростью размножения, измеряли зависимость между плотностью распределения пищи (мух) и количеством пищи, съеденной в единицу времени одной особью. Данные представлены в таблице 3.3 и на рис.3.6.
| Среднее число мух на 1 м 2 | 0 | 0,001 | 0,002 | 0,004 | 0,007 | 0,012 | 0,017 | 0,02 | 0,03 |
| Среднее число мух, съеденных одной особью | 0 | 5,7 | 9,4 | 14,5 | 18,5 | 21,6 | 23,0 | 23,4 | 25,3 |
Рис.3.6. Зависимость между плотностью распределения пищи и количеством пищи, съеденной в единицу времени, для популяции богомола
Вопросы: Определить по представленным данным кинетические параметры интенсивности поедания пищи богомолом, а также плотность распределения пищи, мух, при которой скорость размножения популяции богомола будет равна половине от максимальной.
Решение: Для ответа на первый вопрос отметим, что судя по графику на рис.3.6. кинетика поедания пищи богомолом подчиняется уравнению типа уравнения Моно:
Искомые параметры уравнения Моно I max и K S определим с помощью метода Корниша-Боудена, как это описано в задаче 3.1., используя данные из таб.3.3. Соответствующее решение предсталено на рис. 3.7.
Таким образом, для кинетики поедания мух богомолом I max = 28.5 усл.ед., K S = 0.0038 мух/м 2 .
Ответ на второй вопрос. Из пропорциональности между интенсивностью поедания пищи I и скоростью размножения r следут, что . Отсюда очевидно, что половинная скорость размножения популяции, 0.5 r ( S ), будет наблюдаться, когда плотность распределения мух S будет равна K S , т.е. 0.0038 мух/м 2 .
Рис.3.7. Определение кинетических параметров I max и K S по данным из таб.3.3 .
Условия: Исследовали кинетику размножения оленьей мыши исходя из тех же соображений, что и в задаче 3.2. Полученные данные о зависимости между плотностью распределения пищи, среднего числа коконов пильщиков на 1 м 2 , и средним числом коконов, съеденных одной мышью, представлены в таблице 3.4. и на рис. 3.8.
Среднее число коконов пильщиков на 1 м 2
Вопросы: Проанализировать кинетику поедания коконов пильщиков оленьей мышью. Определить значения соответствующих параметров.
Рис.3.8. Зависимость между средним числом коконов пильщиков на 1 м 2 и средним числом коконов, съеденных одной мышью
Решение: На первом этапе анализа обратим внимание на то, что кривая зависимости между средним числом коконов пильщиков на 1 м 2 и средним числом коконов, съеденных одной мышью, представленная на рис.3.8, имеет два явно отличающихся участка. Первый начинается от нуля и заканчивается примерно на значении 40 коконов на 1 м 2 и характеризуется низкими значениями интенсивности поедания. Далее идет второй участок с существенно более высокими значениями интенсивности поедания, переход одного участка в другой характеризуется четко выраженным изломом на кривой.
Из этого наблюдения следует, что оленья мышь использует более сложный тип пищевого поведения. При низких плотностях пищи очевидно тип питания неспецифичен и характеризуется низким сродством к данному виду пищи. Когда же плотность пищи превышает некоторый порог, тип питания становится специфичным с повышением и сродства и скорости поедания.
Для проверки данного предположения построим график Корниша-Боудена, используя данные из таблицы 3.4.
Видно, что точки пересечения прямых на рис.3.9. группируются в двух различных областях. Это подтверждает предположение о двух кинетических типах пищевого поведения оленьей мыши: первый тип характеризуется высоким значением константы насыщения (низким сродством к пище) и низким значением максимальной интенсивности поедания пищи, второй тип – наоборот.
Рис.3.9. График Корниша-Боудена для данных из таб.3.4.
Переход от типа к типу наблюдается при плотности пищи примерно 40-50 коконов на м 2 (см рис.3.8). Для определения более точных значений кинетических параметров построим графики Корниша-Боудена отдельно для каждого участка (каждого типа пищевого поведения). Очевидно, что для второго участка с высокоспецифичным типом пищевого поведения значения абсцисс нужно брать с поправкой, уменьшать на 40 единиц. Рисунки 3.10 и 3.11 демонстрируют данный подход.
Таким образом, кинетика поедания коконов пильщиков оленьей мышью характеризуется наличием двух участков на оси плотности пищи: на первом участке (0-45 коконов на м 2 ) кинетика низкоспецифична и имеет параметры K S 1 = 250, I max 1 = 70, на втором участке (> 50 коконов на м 2 ) она более специфична с параметрами K S 2 = 7 0, I max 2 = 110.
Рис.3.10 и 3.11. Нахождение кинетических параметров I max и K S для двух различных участков пищевого поведения оленьей мыши.
K S 1 = 250, I max 1 = 70; K S 2 = 7 0, I max 2 = 110;
Основные фазы роста микроорганизмов

ГЛАВА 8. РОСТ МИКРООРГАНИЗМОВ
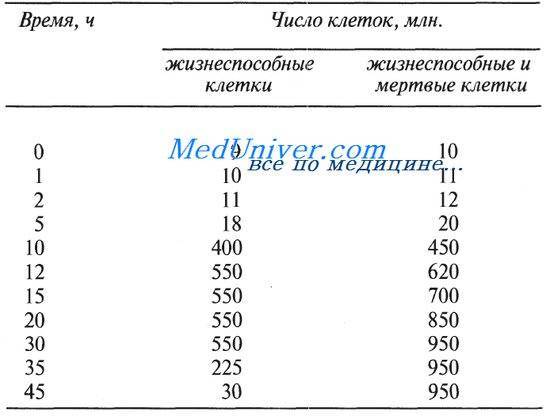
Обмен веществ в микробной клетке сопровождается биосинтезом белков, нуклеиновых кислот, полисахаридов и других соединений. В результате этих процессов увеличиваются размеры и биомасса клетки, что определяется как рост. Увеличение численности клеток в процессе их деления называют размножением. Интенсивность роста определяется путем деления биомассы на численность клеток в единице объема через определенные промежутки времени.
Бактерии, размножающиеся в замкнутом объеме питательной среды, представляют собой микробную популяцию. В лабораторных условиях и на производстве микробные популяции культивируют периодическим или непрерывным методом.
8.1. ПЕРИОДИЧЕСКОЕ КУЛЬТИВИРОВАНИЕ. КРИВАЯ РОСТА
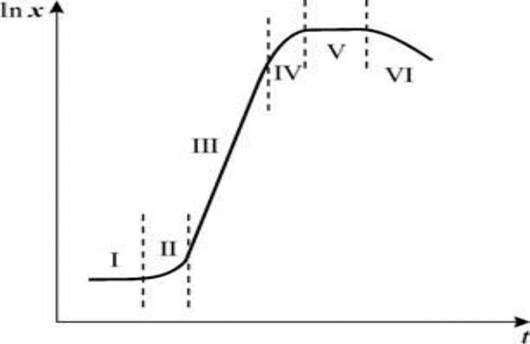
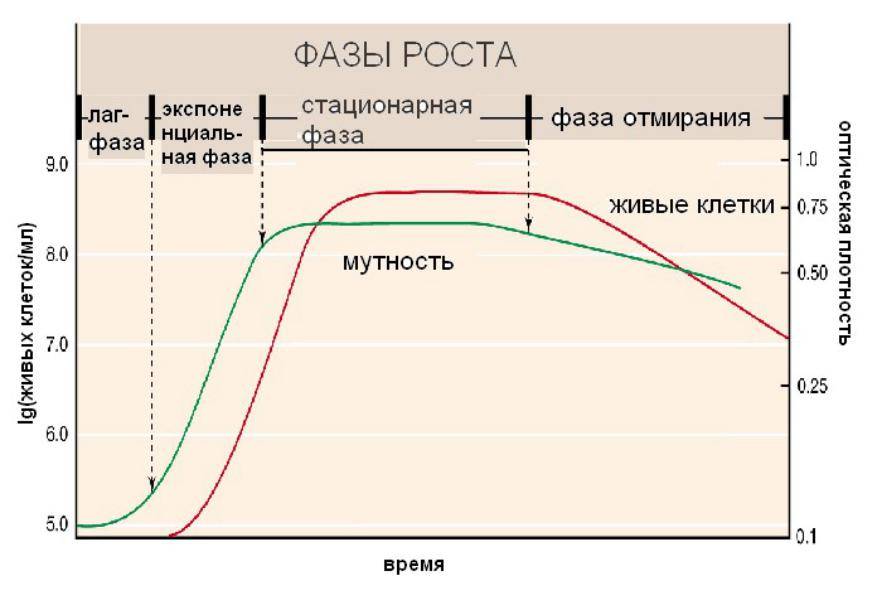
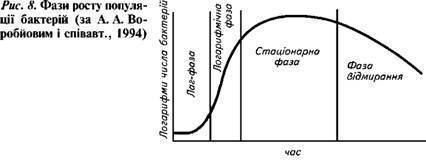
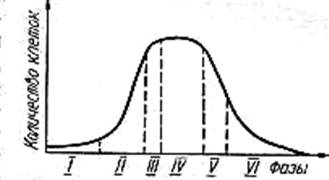
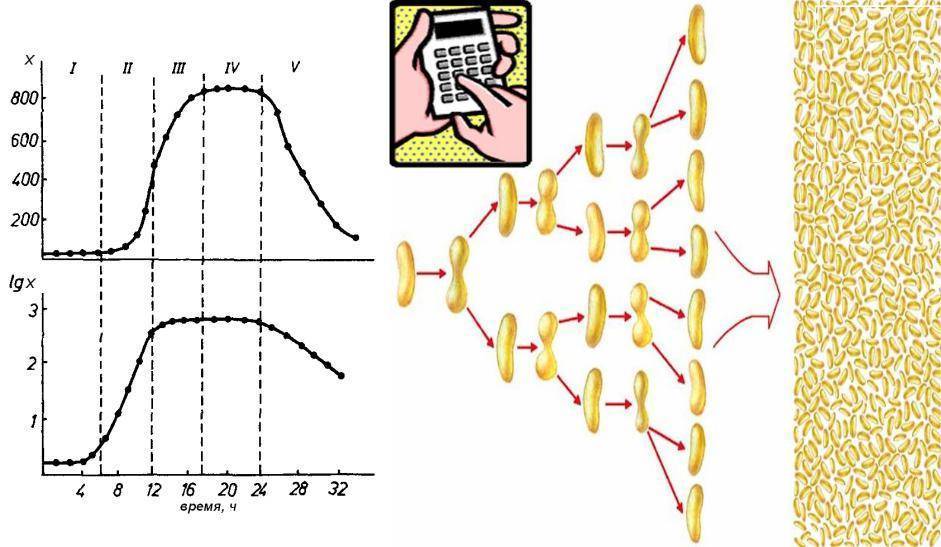
Периодическим культивированием называют размножение микроорганизмов в замкнутом объеме питательной среды. Если в процессе культивирования в среду не добавлять питательные вещества и не выводить продукты жизнедеятельности, то рост микроорганизмов будет продолжаться лишь до тех пор, пока содержание какого-либо компонента не будет исчерпано. Рост микроорганизмов в такой замкнутой системе подчиняется определенным закономерностям и описывается так называемой «кривой роста», выражающей зависимость логарифма числа клеток от времени культивирования (рис. 23).
Рис. 23. Кривая роста микробной популяции в периодических условиях
На типичной кривой роста можно различить несколько фаз роста, сменяющих друг друга в определенной последовательности:
• I — лаг-фаза (или фаза задержки роста);
• II — экспоненциальная (или логарифмическая);
• IV — фаза отмирания.
Лаг-фаза охватывает промежуток времени от момента внесения инокулята (посевного материала) до начала размножения микроорганизмов. Во время лаг-фазы число клеток или совсем не увеличивается, или нарастает очень незначительно. Задержка роста в начале лаг-фазы объясняется адаптацией к новым условиям. Тем не менее этот период характеризуется повышенной метаболической активностью: в клетках возрастает содержание РНК и общего белка, синтезируется ряд ферментов. Наблюдается заметное увеличение размеров клеток — они в 3— 5 раз больше обычных. Длительность лаг-фазы зависит от вида микроорганизма, состава питательной среды, количества и возраста вносимого инокулята. Лаг-фаза сокращается при использовании полноценной среды и молодого инокулята.
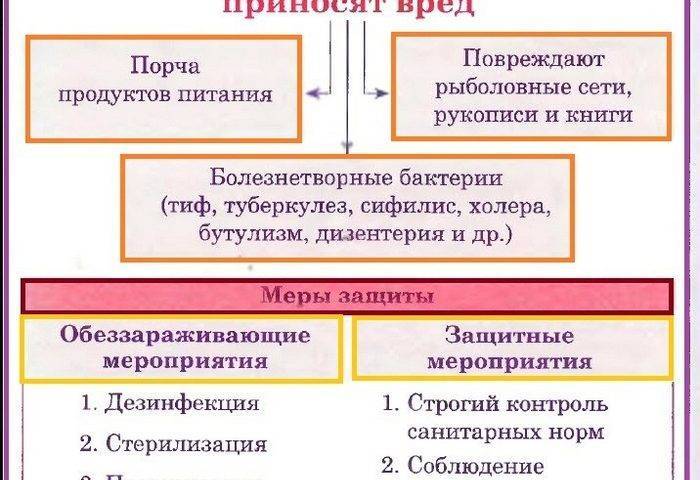

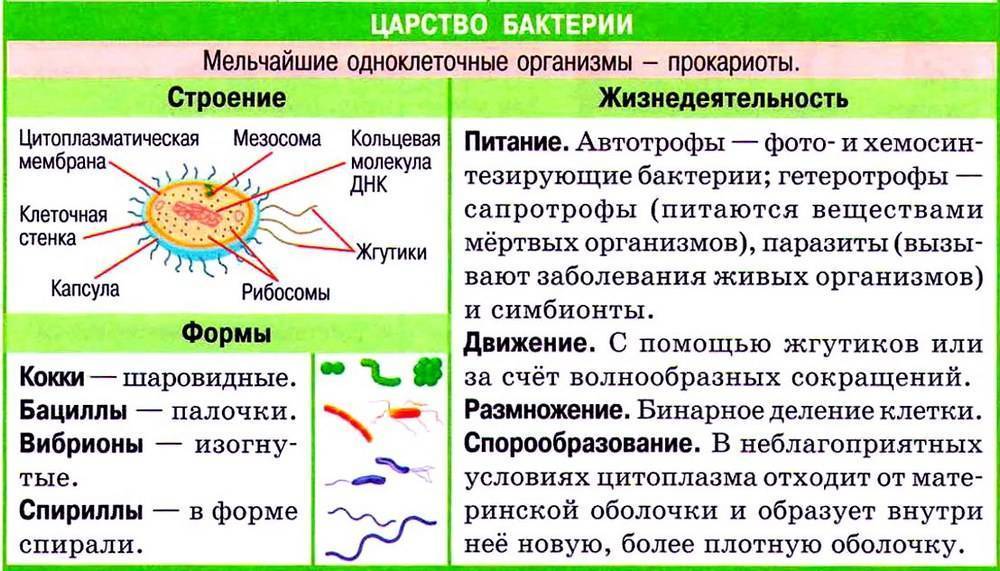

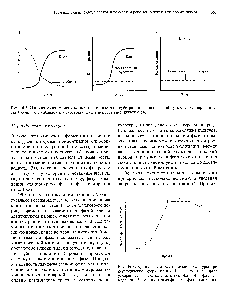

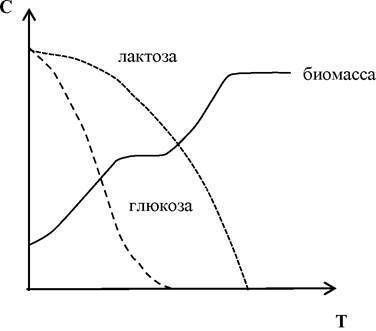

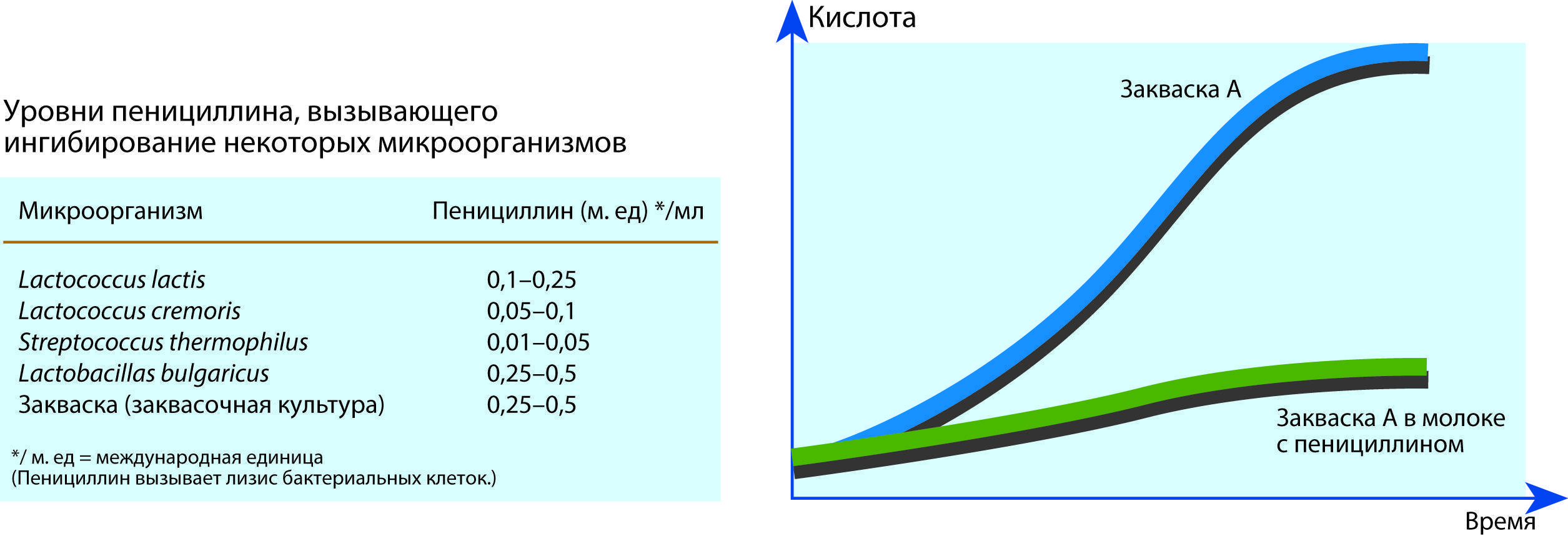
Иногда в периодической культуре может наблюдаться наличие двух лаг-фаз — диауксия. Это явление двухфазного роста в 1942 г. впервые описал французский ученый Жак Моно на примере поочередного использования двух разных углеводов кишечной палочкой. При культивировании кишечной палочки на среде, содержащей глюкозу и сорбит, клетки сначала потребляют глюкозу, тогда как синтез ферментов, необходимых для расщепления сорбита, подавляется. Эти ферменты синтезируются лишь после того, как вся глюкоза будет израсходована.
Экспоненциальная (или логарифмическая) фаза характеризуется максимальной и практически постоянной скоростью роста клеток. Число клеток в этой фазе возрастает в геометрической прогрессии. В этой фазе рост культуры сбалансирован: питательные вещества субстрата и продукты жизнедеятельности не лимитируют размножение микроорганизмов.
В экспоненциальной фазе роста, длительность которой для многих культур микроорганизмов на жидких средах невелика, питательные вещества быстро потребляются, накапливаются вредные продукты обмена, снижается интенсивность синтеза внутриклеточных компонентов. Все это приводит к замедлению роста культуры и переходу ее в следующую фазу — стационарную.
Стационарная фаза соответствует периоду, когда число жизнеспособных клеток, достигнув максимума, перестает увеличиваться. Многие клетки еще продолжают делиться, но количество вновь образующихся клеток равно количеству погибающих. При этом общая численность микробной популяции не изменяется. К концу стационарной фазы условия роста продолжают ухудшаться и культура переходит в завершающую фазу.
Фаза отмирания характеризуется массовой гибелью микробных клеток. Скорость гибели клеток намного опережает скорость их деления. В сложившихся неблагоприятных условиях выживают лишь единичные особи, появляются так называемые инволюционные формы, не дающие потомства.
Особенности периодического культивирования
Различают два основных способа культивирования микроорганизмов — периодическое и непрерывное. При периодическом культивировании клетки помещают в закрытый сосуд определенного объема, содержащий питательную среду, и задают начальные условия. Развитие культуры приводит к постепенной смене условий существования микроорганизмов: увеличивается плотность популяции, снижается концентрация питательных веществ и накапливаются продукты обмена. Периодическую культуру можно рассматривать как замкнутую систему, которая проходит разные фазы своей жизни (рис. 5.5, а).
Рис. 5.5. Типичные кривые роста:
а — при периодическом культивировании; б — диауксия на среде, содержащей два доступных питательных субстрата
Каждая фаза характеризуется определенными физиологическими параметрами. Лаг-фаза — это фаза «привыкания» клеток к среде, при этом происходят увеличение количества ДНК и РНК и индукция синтеза соответствующих ферментов. Лаг-фаза удлиняется, если брать старый посевной материал и переносить клетки в совершенно новую по составу среду. Лаг-фаза сокращается (или может совсем отсутствовать), если активные молодые клетки перенести в свежую среду того же состава и той же температуры. На средах, содержащих смесь субстратов, наблюдается диа- уксия, при которой после исчерпания одного субстрата культура переходит во вторую лаг-фазу для подготовки к потреблению другого субстрата (рис. 5.5, б). В экспоненциальной (логарифмической) фазе клетки растут и делятся с максимальной скоростью, их рост не ограничен. Обычно такие клетки используют в биохимических и физиологических исследованиях. По мере исчерпания субстратов и накопления продуктов обмена скорость роста снижается (наступает фаза замедления роста) и культура переходит в стационарную фазу, в течение которой процессы деления и отмирания клеток в популяции находятся в динамическом равновесии, когда процессы деления и отмирания клеток в популяции уравновешены. Для бактерий эта фаза достигается при концентрации в среднем 109 клеток/мл, для водорослей и простейших — 106 клеток/мл. Когда исчерпание питательных веществ и накопление продуктов метаболизма достигают неких пороговых концентраций, начинается фаза отмирания и число клеток в популяции постепенно снижается. Фаза отмирания обычно тоже имеет логарифмический характер.





Для проведения расчетов р принимались некоторые допущения, при которых скорость изменения плотности клеток в саморазмножающейся системе пропорциональна самой плотности, т.е. изменение в экспоненциальной фазе роста следует кинетике реакций первого порядка. Тогда
Периодическое культивирование
В процессе культивирования микроорганизмов периодическим способом, как указывалось ранее, можно выделить несколько периодов роста (рис. 2.19).

В первый период, после внесения в среду посевного материала (лаг-фаза), происходит процесс приспособления посевной культуры к новой среде. Численность популяции в эта время не увеличивается (а в некоторых случаях даже снижается). Состояние популяции в лаг-фазе формально можно описать так:
(для т, лежащего между 0 и т1).
Предполагается, что в период лаг-фазы микробные клетки не потребляют субстрата, но метаболическая активность клеток проявляется в повышении содержания белка и РНК (при постоянстве содержания ДНК), а также в увеличении объема клеток, который в общем виде может быть выражен с помощью уравнения
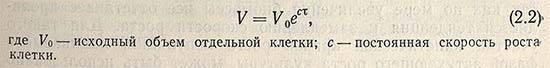
По достижении определенных соотношений между величинами поверхности клетки и ее объема происходит деление клетки, вследствие чего численность популяции начинает увеличиваться с возрастающей скоростью, которая для данной фазы роста культуры, называемой переходной, описывается соотношением
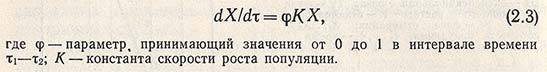
Интегральная зависимость, описывающая участок кинетической кривой роста между т1 и т2, имеет вид
Увеличение скорости роста популяций в переходной фазе идет до предела, определяемого формально достижением параметром ф величины, равной единице, после чего скорость роста начинает выражаться зависимостью
(для т между т2 и т3), откуда интегральная форма представляет экспоненциальную функцию
Эта фаза роста носит название экспоненциальной, или фазы логарифмического роста. Для оценки скорости роста биомассы часто пользуются величиной удельной скорости роста u.



В качестве характеристики растущей культуры, находящейся в этой фазе, используют термины «время удвоения» и «время генерации» q, рассчитываемое по уравнению
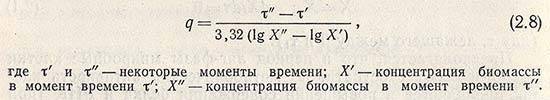
Однако такой характер роста популяции, который в первом приближении может быть описан экспоненциальной зависимостью, наблюдается в течение ограниченного периода времени, так как по мере увеличения биомассы все отчетливее проявляется тенденция к замедлению скорости роста. Для такого участка кинетической кривой роста популяции, называемого фазой затухающего роста культуры, может быть использовано дифференциальное уравнение для скорости роста
и его интегральная форма для описания изменения концентрации биомассы во времени
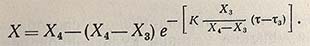
(для интервала времени между т3 и т4).
Снижение скорости роста по мере приближения X к значению Х4 происходит вплоть до достижения нулевого значения, которое характеризует вступление популяции в стационарную фазу: X = X4
(для т, лежащих между т4 и т5).
По завершении фазы стационарного роста начинается фаза отмирания, или фаза дегенерации, культуры, характеризующаяся уменьшением численности популяции.
Приведенная выше система уравнений может быть использована только для описания конкретной кинетической кривой роста, полученной в результате эксперимента, но не в состоянии служить основой для прогнозирования процесса, так как в приведенных зависимостях в качестве параметров (Х1, …, Х4; т1, …, т5) вводятся конечные значения концентрации биомассы и времени. В настоящее время еще нет общепринятой математической модели роста популяции, которая достаточно точно описывала бы кинетику накопления биомассы в условиях периодического культивирования и содержала бы минимальное число эмпирических коэффициентов. В наибольшей степени этим требованиям отвечает модель Н. И. Кобозева, использование которой при изучении кинетики роста популяции дает обнадеживающие результаты. Интегральная форма предложенного им уравнения, описывающего кинетическую кривую роста популяции, имеет вид



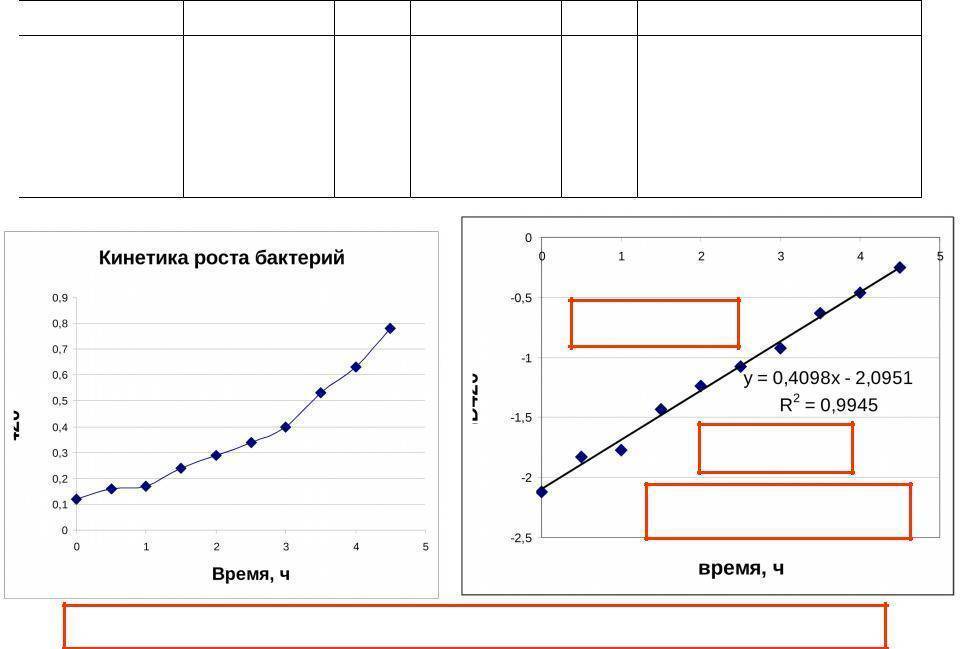



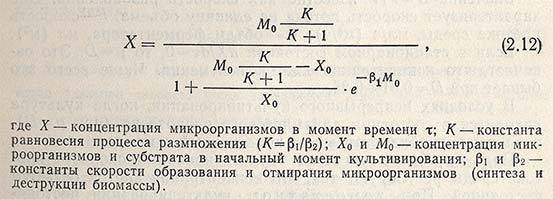
Это уравнение является наиболее общим выражением для роста популяции, и в зависимости от частных условий (обратимое или необратимое размножение, рост популяции с исчерпыванием субстрата или при поддержании его количества на постоянном уровне) уравнение приобретает соответствующую форму и дает различное выражение для величины концентрации биомассы.
Основной недостаток периодического способа — цикличность и постоянное изменение условий культивирования, что затрудняет контроль и регулирование параметров процесса.
Большие возможности для повышения эффективности производства заложены в непрерывном способе выращивания.
Производная функции и ее практическое применение (стр. 4 )
 | Из за большого объема этот материал размещен на нескольких страницах: 1 2 3 4 |

Задача. Функция издержек имеет вид  , а доход при производстве х единиц товара определяется следующим образом:
, а доход при производстве х единиц товара определяется следующим образом:

Определить оптимальное для производителя значение выпуска х0.
Функция прибыли имеет вид:

Найдем производную функции прибыли:

Очевидно, Р'(х)> 0 при х 0 при 100 200, т. е. х=200— максимальное значение Р(х) на интервале (100; + ∞).
Р (200) = 419 900 > Р (100), таким образом, xопт = 200 (ед.).
Задача. Цементный завод производит Х т. цемента в день. По договору он должен ежедневно поставлять строительной фирме не менее 20 т. цемента. Производственные мощности завода таковы, что выпуск цемента не может превышать 90 т. в день.
Определить, при каком объеме производства удельные затраты будут наибольшими (наименьшими), если функция затрат имеет вид:
К=-х3+98х2+200х. Удельные затраты составят К/х=-х2+98х+200
Задача сводится к отысканию наибольшего и наименьшего значения функции
у= — х2+98х+200. На промежутке [20;90].

Вывод: x=49 — критическая точка функции. Вычисляем значение функции на концах промежутках и в критической точке.
f(20)=1760 f(49)=2601 f(90)=320.
Таким образом, при выпуске 49 тонн цемента в день удельные издержки максимальны, это экономически не выгодно, а при выпуске 90 тонн в день минимально, следовательно можно посоветовать работать заводу на предельной мощности и находить возможности усовершенствовать технологию, так как дальше будет действовать закон убывающей доходности. И без реконструкции нельзя будет увеличить выпуск продукции.
6 Применение производной в медицине
Применение дифференциального исчисления в медицине сводится к вычислению скорости. Например, скорости восстановительных реакций и скорости релаксационного процесса.
Реакция организма на введенное лекарство может выражаться в повышении кровяного давления, изменении температуры тела, изменении пульса или других физиологических показателей. Степень реакции зависит от назначенного лекарства, его дозы. С помощью производной можно вычислить, при какой дозе лекарства реакция организма максимальна. С помощью второй производной можно определить условия, при которых скорость процесса наиболее чувствительна к каким-либо воздействиям
Задача Предположим, что х обозначает дозу назначенного лекарства, у — функция степени реакции. у=f(x)=x²(a-x), где а — некоторая положительная постоянная. При каком значении х реакция максимальна?
 .Значит
.Значит  . Тогда
. Тогда  при
при  . В этой точке
. В этой точке  . Значит — тот уровень дозы, который дает максимальную реакцию.
. Значит — тот уровень дозы, который дает максимальную реакцию.
Точки перегиба важны в биохимии, так как они определяют условия, при которых некоторая величина, например скорость процесса, наиболее (или наименее) чувствительна к каким-либо воздействиям.
Задача. В результате значительной потери крови содержание железа в крови уменьшилось на 210 мг. Недостаток железа вследствие его восстановления с течением времени t уменьшается по закону  мг(t – сутки). Найти зависимость скорости восстановления железа в крови от времени. Вычислить эту скорость в момент t=0 и через 7 суток.
мг(t – сутки). Найти зависимость скорости восстановления железа в крови от времени. Вычислить эту скорость в момент t=0 и через 7 суток.
Скорость восстановления железа:

Знак «-» указывает на уменьшение недостачи. При  скорость восстановления равна 30 мг/сутки. Через 7 суток скорость восстановления равна 11,1 мг/сут:
скорость восстановления равна 30 мг/сутки. Через 7 суток скорость восстановления равна 11,1 мг/сут:

Релаксационный процесс – это процесс возвращения системы к состоянию устойчивого равновесия, из которого она была выведена. Во многих случаях (особенно при однократном воздействии) этот процесс описывается экспоненциальным уравнением  , где
, где  – постоянная времени. Ее физический смысл: — это время, в течение которого начальное отклонение
– постоянная времени. Ее физический смысл: — это время, в течение которого начальное отклонение  уменьшается в e раз (т. е. в 2,7 раза). В нашей задаче постоянная времени – 7 суток.
уменьшается в e раз (т. е. в 2,7 раза). В нашей задаче постоянная времени – 7 суток.
7 Применение производной в химии
Скорость химической реакции – один из решающих факторов, который нужно учитывать во многих областях научно-производственной деятельности. Например, инженерам-технологам при определении эффективности химических производств, химикам, разрабатывающим препараты для медицины и сельского хозяйства, а также врачам и агрономам, использующим эти препараты для лечения людей и для внесения их в почву. Одни реакции проходят практически мгновенно, другие идут очень медленно. В реальной жизни для решения производственных задач, в медицинской, сельскохозяйственной и химической промышленности важно знать скорости реакций химических веществ.
Пусть дана функция m=m(t),где m-количество некоторого вещества, вступившего в химическую реакцию в момент времени t. Приращению времени Δt будет соответствовать приращение Δm величины m. Отношение Δm/Δt— есть средняя скорость химической реакции за промежуток времени Δt. Предел этого отношения при стремлении Δt к нулю — есть скорость химической реакции в данный момент времени.
Задача. Зависимость между массой х вещества, получаемого в результате некоторой химической реакции и временем t выражается уравнением  Определите скорость химической реакции в момент времени t.
Определите скорость химической реакции в момент времени t.
Надо найти производную от х по времени t

Задача. Концентрация раствора изменяется с течением времени по закону:  . Найти скорость растворения.
. Найти скорость растворения.
Скорость растворение вычислим с помощью производной:

8 Применение производной в биологии
Биологический смысл производной заключается в том, что по известной зависимости численности популяции можно определить относительный прирост особей.
Пусть зависимость между числом особей популяции микроорганизмов у и временем t её размножения задана уравнением: у=p(t). Пусть Δt-промежуток времени от некоторого начального значения t до t+Δt. Тогда у+Δу=p(t+Δt)— новое значение численности популяции, соответствующее моменту t+Δt, а Δy+p(t+Δt)-p(t)-изменение числа особей организмов.
Задача Рост числа клеток популяции описывается уравнением:  . Получите формулу для скорости роста численности популяции.
. Получите формулу для скорости роста численности популяции.




Задача. Зависимость суточного удоя y в литрах от возраста коров х в годах определяется уравнением  , где х>2. Найдите возраст дойных коров, при котором суточный удой будет наибольшим.
, где х>2. Найдите возраст дойных коров, при котором суточный удой будет наибольшим.

Беря во внимание, что х>2, находим знаки производной на интервалах 

 (лет)- точка максимума, возраст дойных коров, при котором суточный удой будет наибольшим.
(лет)- точка максимума, возраст дойных коров, при котором суточный удой будет наибольшим.
Заключение
В данной работе рассмотрено одно из важнейших понятий математического анализа — производная функции с точки зрения её практического применения. С помощью производной можно решать самые разнообразные задачи, относящиеся к любой области человеческой деятельности. В частности, с помощью производных возможно подробное исследование функций, более точное построение их графиков, решение уравнений и неравенств, доказательство тождеств и неравенств, нахождение наибольших и наименьших значений величин.
По всем вышеперечисленным областям применения производной подобрано и сведено в сборник около двухсот задач. Каждый раздел сборника начинается с краткого изложения теоретических основ, содержит типовые задачи с решениями и наборы упражнений для самостоятельного решения. Эти задачи расширяют кругозор и повышают интерес к производной. Они могут быть интересны и полезны студентам, увлекающимся математикой.
Литература
1. Богомолов задач по математике: учеб пособие для ссузов. – М.: Дрофа, 2005.
2. Богомолов : учеб. для ссузов / , – М.: Дрофа, 2010.
3. Богомолов . Дидактические задания: учеб. пособие для ссузов / , – М.: Дрофа, 2005.
4. Истомина : вопросы и ответы: учеб. пособие для вузов. – Ростов н/Д: Феникс, 2002.
5. Лисичкин : учеб. пособие для техникумов / , — М.:Высш. шк.,1991.
6. Никольский математического анализа: учеб. пособие для студ. ссузов.- М.: Дрофа, 2012.
7. Омельченко : учеб. пособие для ссузов. – Ростов н/Д: Феникс, 2007.
8. Филимонова : учеб. пособие для ссузов. – Ростов н/Д: Феникс, 2013.
http://blotos.ru/rost-i-razmnozenie-bakterij-skorost-i-fazy-rosta-kultivirovanie-bakterij-izbytocnyj-rost
http://pandia.ru/text/80/244/63829-4.php