Синтез холестерина уравнения реакций до мевалоната
Синтез холестерола должен быть согласован с его выведением
Синтез холестерола в организме составляет примерно 0,5-0,8 г/сут, примерно 50% образуется в печени, около 15% в кишечнике. Все клетки организма способны синтезировать холестерол. Поступление с пищей составляет около 0,4 г/сут.
Единственным реальным способом выведения холестерола является желчь – до 1 г/сут.
Биосинтез холестерола
Биосинтез холестерола происходит в эндоплазматическом ретикулуме. Источником всех атомов углерода в молекуле является ацетил-SКоА , поступающий сюда из митохондрий в составе цитрата, также как при синтезе жирных кислот. При биосинтезе холестерола затрачивается 18 молекул АТФ и 13 молекул НАДФН.
Образование холестерола идет более чем в 30 реакциях, которые можно сгруппировать в несколько этапов.
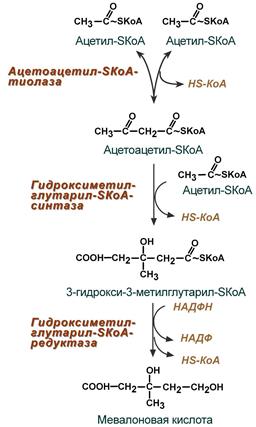
1. Синтез мевалоновой кислоты.
Первые две реакции синтеза совпадают с реакциями кетогенеза, но после синтеза 3-гидрокси-3-метилглутарил-SКоА вступает в действие фермент гидроксиметил-глутарил-SКоА-редуктаза (ГМГ-SКоА-редуктаза), образующая мевалоновую кислоту.
Реакции синтеза мевалоновой кислоты
Схема реакций синтеза холестерола
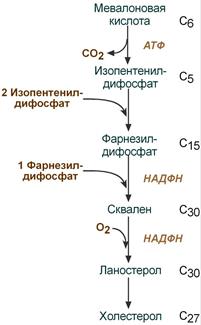
2. Синтез изопентенилдифосфата . На этом этапе при использовании АТФ мевалоновая кислота трижды фосфорилируется. Затем промежуточный продукт декарбоксилируется и дефосфорилируется с получением изопентенилдифосфата.
3. После объединения трех молекул изопентенилдифосфата (если не считать промежуточных превращений) синтезируется фарнезилдифосфат .
4. Синтез сквалена происходит при связывания двух остатков фарнезилдифосфата.
5. После сложных реакций линейный сквален циклизуется в ланостерол .
6. Удаление лишних метильных групп, восстановление и изомеризация молекулы приводит к появлению холестерола .
Регуляция синтеза
Регуляторным ферментом является гидроксиметилглутарил-SКоА-редуктаза , активность которой может изменяться в 100 и более раз.
1. Метаболическая регуляция – по принципу обратной отрицательной связи фермент аллостерически ингибируется конечным продуктом реакции – холестеролом. Это помогает поддерживать внутриклеточное содержание холестерола постоянным.
2. Регуляция транскрипции гена ГМГ-SКоА-редуктазы (в печени) – холестерол и желчные кислоты подавляют считывание гена и уменьшают количество фермента.
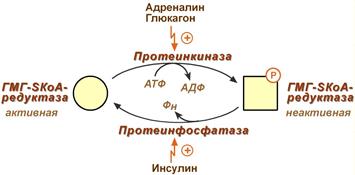
3. Ковалентная модификация при гормональной регуляции:
Инсулин, активируя протеин-фосфатазу, способствует переходу фермента в активное состояние.
Глюкагон и адреналин посредством аденилатциклазного механизма активируют протеинкиназу А, которая фосфорилирует фермент и переводит его в неактивную форму.
Регуляция активности гидроксиметилглутарил-S-КоА-редуктазы
Кроме указанных гормонов, на ГМГ-SКоА-редуктазу действуют эстрогены и тиреоидные гормоны (повышают активность) и глюкокортикоиды (снижают активность).
Изменение транскрипции гена ГМГ-КоА редуктазы (генетическая регуляция) осуществляется стерол-регулируемым элементом в ДНК (SREBP, sterol regulatory element-binding protein) с которым способны связываться белки — факторы SREBP. Эти факторы при достаточном количестве холестерина в клетке закреплены в мембране ЭПР. Когда уровень холестерина падает, факторы SREBP активируются под действием специфичных протеаз комплекса Гольджи, передвигаются в ядро, взаимодействуют на ДНК с участком SREBP и стимулируют биосинтез холестерина.
Скорость биосинтеза холестерола также зависит от концентрации специфического белка-переносчика, обеспечивающего связывание и транспорт гидрофобных промежуточных метаболитов синтеза.
Биосинтез холестерина: реакции до мевалоновой кислоты далее, схематично.
В синтезе холестерина можно выделить три основные стадии: I – превращение активного ацетата в мевалоновую кислоту, II – образование сквалена из мевалоновой кислоты, III – циклизация сквалена вхолестерин.
I этап синтеза ХС — образование мевалоната (мевалоновой кислоты).
1. Две молекулы ацетил-КоА конденсируются тиолазой с образованием ацетоацетил-КоА;
2. Гидроксиметилглутарил-КоА-синтаза присоединяет третий ацетильный остаток к ацетоацетил-КоА с образованием ГМГ-КоА (3-гидрокси-3-метилглутарил-КоА). Эта последовательность реакций сходна с начальными стадиями синтеза КТ. Однако синтез КТ происходит в митохондриях печени, а реакции синтеза ХС — в цитозоле клеток.
3. ГМГ-КоА-редуктаза восстанавливает ГМГ-КоА до мевалоната с использованием 2 молекул НАДФH2. Фермент ГМГ-КоА-редуктаза — гликопротеин, пронизывающий мембрану ЭПР, активный центр которого выступает в цитозоль.
II этап синтеза ХС — образование сквалена
1. Мевалонат превращается в изопреноидную структуру — изопентенилпирофосфат (5 атомов С);
2. 2 изопентенилпирофосфата конденсируются в геранилпирофосфат (10 атомов С);
3. Присоединение изопентенилпирофосфата к геранилпирофосфату дает фарнезилпирофосфат (15 атомов С).
4. 2 фарнезилпирофосфата конденсируются в сквален (15 атомов С).
III этап синтеза ХС — образование ХС
Рассмотрим стадию превращения активного ацетата в мевалоновую кислоту. Начальным этапом синтеза мевалоновой кислоты из ацетил-КоА является образование ацетоацетил-КоА посредством обратимой тиолазной реакции:
Затем при последующей конденсации ацетоацетил-КоА с 3-й молекулой ацетил-КоА при участии гидроксиметилглутарил-КоА-синтазы (ГМГ-КоА-синтаза) образуется β-гидрокси-β-метилглутарил-КоА:
Далее β-гидрокси-β-метилглутарил-КоА под действием регуляторно-го фермента НАДФ-зависимой гидроксиметилглутарил-КоА-редуктазы (ГМГ-КоА-редуктаза) в результате восстановления одной из карбоксильных групп и отщепления HS-KoA превращается в мевалоновую кислоту:
Далее общая картинка,откуда можно просто схематично запомнить))
Описанный путь биосинтеза локализован в гладком ЭР. Синтез идет за счет энергии, освобождающейся при расщеплении производных коферментаА и энергетически богатых фосфатов. Восстановителем при образовании мевалоната и сквалена, а также на последних стадиях биосинтеза холестерина является НАДФН + Η + . Для этого пути характерно то, что промежуточные метаболиты можно подразделить на три группы: производные кофермента А, дифосфаты и высоко липофильные соединения (от сквалена до холестерина), связанные с переносчиками стеринов.
43.Особенности регуляции в кишечной стенке и других тканях биосинтеза ХС; роль гормонов: инсулина, Т3,Т4, витамина РР.
Регуляция синтеза ХС
Ключевой фермент синтеза ХС ГМГ-КоА-редуктаза регулируется несколькими способами:
В печени синтез редуктазы подвержен суточным колебаниям: максимум на полночь и минимум на утренние часы.
· ХС, желчные кислоты (в печени) репрессируют ген ГМГ-КоА-редуктазы. В норме поступление ХС с пищей снижает синтез собственного ХС в печени, однако с возрастом эффективность этой регуляции у многих людей снижается и уровень ХС повышается.
· Инсулин через дефосфорилирование осуществляет активацию ГМГ-КоА-редуктазы.
· Глюкагон через фосфорилирование осуществляет ингибирование ГМГ-КоА-редуктазы.
Повышение концентрации исходного субстрата ацетил-КоА стимулирует синтез ХС.
Таким образом, синтез ХС активируется при питании углеводами и ингибируется при голодании.
Основная роль витамина РР в организме – участие в окислительно-восстановительных процессах. Витамин РР способствует нормальному росту тканей, оказывает благотворное влияние на жировой обмен, участвует в преобразовании сахара и жиров в энергию, снижает в крови уровень «плохого» холестерина.
Активность ГМГ-редуктазы возрастает при введении инсулина и тиреоидных гормонов(Т3 и Т4), это приводит к усилению синтеза ХС.
При голодании, тиреоидэктомии, введении глюкагона и глюкокортикоидов – угнетение синтеза ХС.
ОСНОВЫ БИОХИМИИ ЛЕНИНДЖЕРА — ТОМ 2. БИОЭНЕРГЕТИКА И МЕТАБОЛИЗМ — 2014
ЧАСТЬ II. БИОЭНЕРГЕТИКА И МЕТАБОЛИЗМ
21. БИОСИНТЕЗ ЛИПИДОВ
21.4. Биосинтез холестерина, стероидов и изопреноидов
Без сомнения, холестерин — наиболее известный широкой публике липид; он пользуется дурной славой из-за высокой корреляции между повышенным уровнем холестерина в крови и частотой сердечно-сосудистых заболеваний человека. Меньшее внимание привлечено к решающей роли холестерина как компонента клеточных мембран и как предшественника стероидных гормонов и желчных кислот. Холестерин необходим многим животным, в том числе человеку, но его наличие в пище млекопитающих необязательно — клетки организма сами могут синтезировать его из простых предшественников.
Структура этого 27-углеродного соединения дает основание предположить сложный путь его биосинтеза, однако все его углеродные атомы предоставлены единственным предшественником — ацетатом. Изопреновые блоки — самые главные интермедиаты на пути от ацетата до холестерина, они являются предшественниками многих природных липидов, и механизмы, по которым полимеризуются изопреновые блоки, похожи во всех метаболических путях.
Начнем с рассмотрения главных стадий на пути биосинтеза холестерина из ацетата, затем обсудим транспорт холестерина по кровотоку, поглощение его клетками, нормальную регуляцию синтеза холестерина и регуляцию при нарушениях его захвата или транспорта. Затем мы рассмотрим другие вещества, которые происходят от холестерина, такие как желчные кислоты и стероидные гормоны. Наконец, описание биосинтетических путей образования многих соединений — производных изопреновых блоков, в которых есть общие ранние стадии с синтезом холестерина, проиллюстрирует необычайную универсальность изопреноидной конденсации в биосинтезе.
Холестерин производится из ацетил-СоА в четыре стадии
Холестерин, подобно длинноцепочечным жирным кислотам, производится из ацетил-СоА, но схема сборки совершенно иная. В первых опытах в корм животных добавляли ацетат, меченный 14 С либо по метильному, либо по карбоксильному атому углерода. На основе распределения метки в холестерине, выделенном из двух групп животных (рис. 21-32), были описаны ферментативные стадии биосинтеза холестерина.
Рис. 21-32. Источник углеродных атомов холестерина. Выявлено в ходе экспериментов с использованием радиоактивного ацетата с меткой по метильному углероду (черный цвет) или углероду карбоксила (красный). В конденсированной структуре кольца обозначены буквами от А до D.
Синтез происходит в четыре стадии, как показано на рис. 21-33: (1) конденсация трех ацетатных остатков с образованием шестиуглеродного интермедиата мевалоната; (2) превращение мевалоната в активированные изопреновые блоки; (3) полимеризация шести пятиуглеродных изопреновых единиц с образованием 30-углеродного линейного сквалена; (4) циклизация сквалена с образованием четырех колец стероидного ядра, после чего следует серия изменений (окисление, удаление или миграция метальных групп) с образованием холестерина.
Рис. 21-33. Обобщенная картина биосинтеза холестерина. Четыре стадии синтеза обсуждаются в тексте. Изопреновые блоки в сквалене отмечены красными штриховыми линиями.
Стадия (1). Синтез мевалоната из ацетата. Первая стадия биосинтеза холестерина приводит к образованию промежуточного продукта мевалоната (рис. 21-34). Две молекулы ацетил-СоА конденсируются, давая ацетоацетил-СоА, который конденсируется с третьей молекулой ацетил-СоА с образованием шестиуглеродного соединения β-гидрокси-β-метилглутарил-СоА (НМG-СоА). Эти две первые реакции катализируются тиолазой и НМG-СоА-синтазой соответственно. Цитозольная НМG-СоА-синтаза этого метаболического пути отличается от митохондриального изофермента, который катализирует синтез НМG-СоА при образовании кетоновых тел (см. рис. 17-18).
Рис. 21-34. Образование мевалоната из ацетил-СоА. Источник С-1 и С-2 мевалоната из ацетил-СоА выделен розовым цветом.
Третья реакция лимитирует скорость всего процесса. В ней происходит восстановление НМG-СоА до мевалоната, для которого каждая из двух молекул NАDРН предоставляет два электрона. HMG-СоА-редуктаза — интегральный мембранный белок гладкого ЭР; он служит, как мы увидим далее, главным пунктом регуляции метаболического пути образования холестерина.
Стадия (2). Превращение мевалоната в два активированных изопрена. На следующей стадии синтеза холестерина с молекул АТР на мевалонат переносятся три фосфатные группы (рис. 21-35). Фосфат, связанный с гидроксильной группой при С-3 мевалоната в интермедиате 3-фосфо-5- пирофосфомевалонате, является хорошей уходящей группой; на следующей стадии уходят оба этих фосфата и прилежащая карбоксильная группа, образуя двойную связь в пятиуглеродном продукте ∆ 3 —изопентенилпирофосфате. Это первый из двух активированных изопренов — главных участников синтеза холестерина. Изомеризация ∆ 3 -изопентенилпирофосфата дает второй активированный изопрен диметилаллилпирофосфат. Синтез изопентенилпирофосфата в цитоплазме растительных клеток происходит по описанному здесь пути. Однако хлоропласты растений и многие бактерии используют независимый от мевалоната путь. Этот альтернативный путь не встречается у животных, поэтому он привлекателен при создании новых антибиотиков.
Рис. 21-35. Превращение мевалоната в активированные изопреновые блоки. Шесть активированных единиц объединяются, образуя сквален (см. рис. 21-36). Уходящие группы 3-фосфо-5-пирофосфомевалоната выделены розовым. В квадратных скобках — гипотетический интермедиат.
Стадия (3). Конденсация шести активированных изопреновых единиц с образованием сквалена. Изопентенилпирофосфат и диметилаллилпирофосфат претерпевают теперь конденсацию «голова к хвосту», при которой одна пирофосфатная группа перемещается и образуется 10-углеродная цепь — геранилпирофосфат (рис. 21-36). (Пирофосфат присоединяется к «голове».) Геранилпирофосфат подвергается следующей конденсации «голова к хвосту» с изопентенилпирофосфатом, при этом образуется 15-углеродный интермедиат фарнезилпирофосфат. Наконец, две молекулы фарнезилпирофосфата соединяются «голова к голове», обе фосфатные группы удаляются — образуется сквален.
Рис. 21-36. Образование сквалена. Структура сквалена, содержащая 30 атомов углерода, возникает при последовательных конденсациях, активированных изопреновых (пятиуглеродных) блоков.
Общепринятые названия этих промежуточных продуктов происходят от названий тех источников, откуда они были впервые выделены. Гераниол, компонент розового масла, имеет аромат герани, а фарнезол, найденный в цветах акации Фарнеза, — аромат ландыша. Многие природные запахи растительного происхождения принадлежат соединениям, построенным из изопреновых блоков. Сквален, впервые выделенный из печени акулы (вида Squalus), состоит из 30 атомов углерода: 24 атома в главной цепи и шесть атомов в метальных заместителях.
Стадия (4). Превращение сквалена в четыре кольца стероидного ядра. На рис. 21-37 ясно видно, что у сквалена цепочечная структура, а у стеринов — циклическая. Все стерины имеют четыре конденсированных кольца, которые образуют стероидное ядро, и все они — спирты с гидроксильной группой при атоме С-3; отсюда английское название sterol. При действии скваленмонооксигеназы к концу цепи сквалена добавляется один атом кислорода из O2 и образуется эпоксид. Этот фермент представляет собой еще одну оксидазу со смешанной функцией (доп. 21-1); NADPH восстанавливает другой кислородный атом из O2 до Н2O. Двойные связи продукта сквален-2,3-эпоксида расположены таким образом, что замечательно согласованная реакция может превратить цепочечный сквален- эпоксид в циклическую структуру. В животных клетках эта циклизация приводит к образованию ланостерина, который содержит четыре кольца, характерные для стероидного ядра. В итоге ланостерин превращается в холестерин посредством серии примерно из 20 реакций, которая включает миграцию одних метальных групп и удаление других. Описание этого удивительного пути биосинтеза, одного из самых сложных среди известных, было сделано Конрадом Блохом, Феодором Линеном, Джоном Корнфортом и Джорджем Попьяком в конце 1950-х гг.
Рис. 21-37. Замыкание кольца превращает линейный сквален в конденсированное стероидное ядро. Первая стадия катализируется оксидазой со смешанной функцией (монооксигеназой), косубстрат которой — NADPH. Продукт — эпоксид, который на следующей стадии циклизуется с образованием стероидного ядра. Конечный продукт этих реакций в животных клетках — холестерин; у других организмов образуются слегка отличные от него стерины.
Холестерин — стерин, характерный для животных клеток; растения, грибы и протисты производят другие очень похожие стерины.
Они используют тот же самый путь синтеза до сквален-2,3-эпоксида, но затем пути слегка расходятся, при этом образуются другие стерины, такие как сигмастерин у многих растений и эргостерин у грибов (рис. 21-37).
Пример 21-1 Энергетические затраты при синтезе сквалена
Каковы энергетические затраты (выраженные в числе молекул АТР) на синтез одной молекулы сквалена?
Решение. При синтезе сквалена из ацетил-СоА АТР затрачивается лишь на той стадии, когда мевалонат превращается в активированный изопреновый предшественник сквалена. Для построения молекулы сквалена нужно шесть активированных молекул изопрена, а для получения каждой активированной молекулы требуется три молекулы АТР. Итого, на синтез одной молекулы сквалена затрачивается 18 молекул АТР.
Соединения холестерина в организме
У позвоночных большие количества холестерина синтезируются в печени. Некоторая часть синтезированного там холестерина включается в мембраны гепатоцитов, но в основном он экспортируется в виде одной из трех своих форм: билиарного (желчного) холестерина, желчных кислот или эфиров холестерина. Желчные кислоты и их соли — гидрофильные производные холестерина, которые синтезируются в печени и способствуют перевариванию липидов (см. рис. 17-1). Эфиры холестерина образуются в печени при действии ацил-СоА-холестерин- ацилтрансферазы (АСАТ). Этот фермент катализирует перенос остатка жирной кислоты от кофермента А к гидроксильной группе холестерина (рис. 21-38), превращая холестерин в более гидрофобную форму. Эфиры холестерина в секретируемых липопротеиновых частицах транспортируются в другие ткани, использующие холестерин, или хранятся в печени.
Рис. 21-38. Синтез эфиров холестерина. Этерификация превращает холестерин в еще более гидрофобную форму для хранения и транспорта.
Холестерин необходим всем тканям растущего организма животных для синтеза мембран, а некоторые органы (например, надпочечники и половые железы) используют холестерин в качестве предшественника стероидных гормонов (об этом речь пойдет ниже). Холестерин также является предшественником витамина D (см. рис. 10-20, т. 1).
Холестерин и другие липиды переносят липопротеины плазмы крови
Холестерин и эфиры холестерина, подобно триацилглицеринам и фосфолипидам, практически нерастворимы в воде, тем не менее они должны перемещаться от ткани, в которой были синтезированы, к тканям, где они будут сохраняться или потребляться. Они переносятся кровотоком в виде липопротеинов плазмы крови — макромолекулярных комплексов специфических белков- переносчиков (аполипопротеинов) с фосфолипидами, холестерином, эфирами холестерина и триацилглицеринами, присутствующими в этих комплексах в различной комбинации.
Аполипопротеины («апо» обозначает сам белок без липида) соединяются с липидами, образуя несколько фракций липопротеиновых частиц — сферических комплексов с гидрофобными липидами в центре и гидрофильными аминокислотными цепями на поверхности (рис. 21-39, а). При различных комбинациях липидов и белков образуются частицы разной плотности — от хиломикронов до липопротеинов высокой плотности. Эти частицы можно разделить ультрацентрифугированием (табл. 21-1) и визуально наблюдать с помощью электронной микроскопии (рис. 21-39, б). Каждая фракция липопротеинов выполняет специфическую функцию, которая определяется местом синтеза, составом липидов и содержанием аполипопротеина. В плазме крови человека обнаружены по крайней мере 10 различных аполипопротеинов (табл. 21-2), различающихся по размеру, реакциям со специфическими антителами и характерному распределению в липопротеинах разных классов. Эти белковые компоненты работают как сигнальные вещества, направляющие липопротеины к специфическим тканям или активирующие ферменты, которые действуют на липопротеины.