Математическая модель мышечного сокращения
Высшая Школа Естественных наук и технологий
Кафедра фундаментальной и прикладной физики
Лаборатория биофизики
Указания к лабораторной работе
«Изучение работы мышц»

Архангельск
2018
Теоретическое введение
Биологическая подвижность
Способность к движению – одно из характерных свойств всех живых организмов, начиная от простейших и кончая самыми сложными. Все проявления двигательной активности имеют общую черту – превращение химической энергии, освобождающейся при гидролизе АТФ, в механическую. Белковые структуры, участвующие в гидролизе АТФ и генерации силы — это либо миозин и актин, либо кинезин (или динесен) и тубулин. При мышечном сокращении механическая работа осуществляется организованными в надмолекулярные структуры ферментом – АТФазой миозином и актином. Регулятором двигательной активности в мышцах является кальций (Ca ++ ). Определенную роль в регуляции двигательной активности играют ионы магния Mg ++ .
Режимы сокращения мышц
Термин «сокращение» употребляется к мышце в двух смыслах. Изометрическое сокращение происходит в мышце при её фиксированной длине, т.е. развивается напряжение без укорочения. Изотоническое сокращение означает укорочение мышцы при постоянной нагрузке. В этом случае мышца производит работу, поднимая груз.
Аксотонический режим характеризуется одновременным изменением длины и напряжения в мышце.
Временная зависимость характеристик одиночного сокращения при изометрическом и изотоническом режимах представлена на рис. 1. а, б.

а – изометрический режим б – изотонический режим
Рис. 1.Временная зависимость характеристик при работе мышц
Изометрический режим характеризуется максимальным значением Р0 изометрической силы.
Зависимость ε(t) в изотоническом режиме свидетельствует о том, что мышца является вязкоупругим элементом.
Для вязкоупругой среды напряжение σ = F/S, будет определяться упругой σУ и вязкой σВ составляющими:
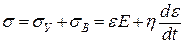 , (1)
, (1)
где Е – модуль Юнга, η – коэффициент вязкости.
Решив дифференциальное уравнение (1), получим:
 , (2)
, (2)
где  называется временем релаксации.
называется временем релаксации.
Зависимость ε(t) на рис. 1б. на участке возрастания близка к зависимости, описываемой формулой (2).
Уравнение Хилла
Экспериментальная методика изменений зависимости скорости изотонического сокращения от нагрузки, измерении напряжения, развиваемого при изометрических сокращениях, а так же методика калориметрических измерений теплообмена мышцы при сокращении была развита в классических работах А.В. Хилла в период с 1938 г. по 1964 г., при исследовании напряжений мышцы лягушки.
Соотношение между основными механическими параметрами мышечного сокращения – скоростью укорочения и развиваемой силой – носит название характеристического уравнения Хилла. Оно имеет вид:
 , (3)
, (3)
где Р0 – изометрическая сила (Р = Р0 при υ = 0), a и b — эмпирические параметры, связанные соотношением:
 (4)
(4)
График зависимости (3) представлен на рис. 2.

Рис. 2.График зависимости по уравнению Хилла
Исследования теплопродукции мышцы при изотонических укорочениях, проведенные А.В. Хиллом, позволили установить, что количество тепла, выделяющегося в сокращающейся мышце, дополнительно к изометрическому теплу, зависит только от величины укорочения:
 (5)
(5)
Тогда скорость теплопродукции dQ/dt пропорциональна скорости укорочения  . При укорочении мышца совершает работу
. При укорочении мышца совершает работу  .
.
Тогда получим для скорости избыточной энергопродукции:
 (6)
(6)
Уравнение (6) является вторым уравнением Хилла.
Эксперименты показали что коэффициент А зависит от нагрузки:
 (7)
(7)
Следует отметить в изометрическом режиме происходит генерация изометрического тепла:
 (8)
(8)
График зависимости (6)представлен на рис. 3.

Рис. 3.Зависимость – уравнение Хилла
На рис. 4. представлена зависимость механической мощности от нагрузки.

Рис. 4.Зависимость механической мощности от нагрузки
По количеству выделяемой мышцей тепла можно оценить коэффициент полезного действия мышцы. При Р = (0,2 ÷ 0,8) Р0 К.П.Д. составляет около 40%.
Модель скользящих нитей
Мышечная ткани представляет собой совокупность мышечных волокон, внеклеточного вещества и густой сети нервных волокон и кровеносных сосудов.
Отдельное мышечное волокно имеет диаметр 20 – 80 мкм, и окружено плазматической мембраной толщиной 10 нм. Каждое волокно – это сильно вытянутая клетка. Длина отдельных волокон может существенно варьироваться, в зависимости от вида мышцы от сотен микро до нескольких сантиметров.
Сокращательный аппарат клетки состоит из 1000 – 2000 параллельно расположенных миофибрилл диаметром 1 – 2 мкм, В миофибриллах различают: А – зону, темные полосы, которые в поляризованном свете дают двойное лучепреломление, I – зону – светлые полосы, не дающие двойного лучепреломления. В области I – зоны наблюдается темная узкая полоса Z – диск (смотри рис. 5)

Рис. 5.Сократительный аппарат клетки
Промежуток между Z – дисками называется саркомером и является элементарной сократительной единицей. Макроструктура саркомера представлена на рис. 6

Рис. 6.Макроструктура саркомера
Саркомер – упорядоченная структура толстых и тонких нитей, расположенных гексагонально в поперечном сечении. Толстая нить имеет толщину 12 нм и длину 1,5 мкм и состоит из белка миозина. Тонкая нить имеет диаметр 8 нм и длину 1 мкм и состоит из белка актина, прикрепленного одним концом к Z – диску. Актиновая нить состоит из двух закрученных один вокруг другого мономеров актина диаметром 5 нм, что похоже на две нитки бус скрученных по 14 бусин в витке. В цепях актина через 40 нм встроены молекулы тропонина.
При сокращении мышцы тонкие нити вдвигаются между толстыми. Происходит относительное скольжение нитей без изменения их длин. Этот процесс обусловлен взаимодействием особых выступов миозина – поперечных мостиков с активными центрами, расположенными на актине. Мостики отходят от толстой нити периодично при расстоянии 14,5 нм.
В расслабленном состоянии миофибрилл молекулы тропомиозина блокируют прикрепление поперечных мостиков. При подаче стоимулирующего импульса ионы Ca ++ активизируют мостики и открывают участки их прикрепления к актину (рис.7).

Рис. 7.Процесс работы актиновых нитей
В момент прикрепления происходит расщепление АТФ, вследствие гидратации. Это приводит к повороту головки мостика. В результате этого процесса происходит генерация силы, скольжение актина относительно миозина к центру саркомера и как следствие – укорочение мышцы.
Таким образом актин-миозиновый комплекс является механо-химическим преобразователем энергии, выделяющейся при гидратации АТФ.
Математическая модель мышечного сокращения
Наиболее простая математическая модель мышечного сокращения была предложена В.И. Дещеревским в 1968 г. на основе моделей скользящих нитей.
Скорость изменения длинны волокна  , где N – число саркомеров в волокне,
, где N – число саркомеров в волокне,  — относительная скорость скольжения нитей.
— относительная скорость скольжения нитей.
Кинетическая схема переходов мостика между различными состояниями представлена трехстадийным принципом:

Выделяется три состояния: замкнутое, замкнутое тянущее (головка генерирует силу +f), замкнутое тормозящее (головка генерирует тормозящую силу –f). На схеме δ — длинна зоны, в которой мостик развивает тянущую силу, К1 – константа скорости замыкания свободного мостика, К2 – константа распада тормозящих мостиков.
Сила развиваемая саркомером: F = f·n ‑ f·m. В соответствии с циклом для числа тянущих (n) и тормозящих (m) мостиков можно записать систему дифференциальных уравнений:
 (9)
(9)
 (10)
(10)
где α0 – полное число доступных для замыкания мостиков.
При стационарном режиме сокращения, когда υ = const, производные от n и m равны нулю, а сумма сил, развиваемых тянущими и тормозящими мостиками, равна нагрузке приложенной к мышце:
Решая систему уравнений (9-11) получаем, для стационарного состояния
 (12a)
(12a)
 (12б)
(12б)
 (12в)
(12в)
Следует отметить, что уравнение 12в соответствует феноменологическому уравнению Хилла (3), если положить:

Сопоставляя с экспериментальными результатами для портняжной мышцы лягушки: Р0 = 30 Н/см 2 , a/P0 = 0,25 и υmax = 1,5·10 -6 м/с и, полагая, что в мышечном слое толщиной в половину саркомера с поперечным сечением 1 см 2 число мостиков α0 = 10 13 и энергия гидролиза одной молекулы АТФ ε = 3·10 -20 Дж и полагая, что значение положительной работы в цикле fδ = ε, получим: f = 3·10 -12 H; δ = 10 -6 см; К1 = 50 1/см; К2 = 150 1/см.
Стационарная скорость энергопродукции в кинематической теории пропорциональна скорости размыкания тормозящих мостиков
 (13)
(13)
где ε — энергия освобождающаяся при гидролизе одной молекулы АТФ.
ХИЛЛА УРАВНЕНИЕ
Найдено 20 изображений:
ХИЛЛА УРАВНЕНИЕ мышечного сокращения, выражает изменение скорости сокращения мышцы в зависимости от её нагрузки. Выведено англ, физиологом А. В. Хиллом в 1938. Формула X. у.: (P + a)(v + b) = = о(Ро + а), где v — скорость сокращения мышцы при нагрузке Р, Ро-макс, значение изометрич. силы при тетани-ческом (см. Тетанус) раздражении всей мышцы, константы а и b — эмпирич. величины. Константа а имеет размерность силы и равна ок. 4-10 5 дин/см 2 поперечного сечения мышц различных видов, а константа b имеет размерность скорости (выражается в см/сек или to /сек, где /о — начальная длина мышцы) и для разных мышц различна.
В более общем виде эту закономерность выразили в 1953 англ, учёные Б. С. Эббот и Д. Р. Уилки. Если сокращающаяся мышца имеет длину I в момент времени t, то скорость её укорочения-dl/dt определяется по формуле: -dl/dt = (Ft — F)b/(F + а), где F -сила, к-рую преодолевает мышца, FI -макс, сила мышц при той длине, при к-рой измеряется скорость её укорочения, а и b — константы. Эта формула модифицирована Уилки в 1956, что позволило рассматривать скорость сокращения мышцы ( —dx/dt) при любой заданной нагрузке во время тетанич.
напряжение мышцы, пропорциональное тетанич. раздражению, fi(Fm) —характеристика зависимости напряжения от нагрузки для упругого элемента, соединённого последовательно, FO — изометрич. (тетанич.) напряжение.
Скорость сокращения уменьшается при понижении темп-ры; температурный коэффициент Q10 равен ок. 2,5. Даже при отсутствии силы, противодействующей сокращению, мышца укорачивается с ограниченной скоростью: если F = 0, то -(dl/dt) = F1b/a.
X. у. точно описывает сокращение мышц позвоночных и беспозвоночных, хотя ещё не установлено соответствие констант уравнения сократит., упругим и вязким элементам структуры мышцы. См. также Мышечное сокращение.
Лит.: Физиология мышечной деятельности, труда и спорта, Л., 1969 (Руководство по физиологии); Хилл А., Механика мышечного сокращения, пер. с англ., М., 1972; Abbott В. С., W i I k i е О. R., The relation between velocity of shortening and the tension-length curve of skeletal muscle, «Journal of Physiology», 1953, v. 120; W i 1 k i e D. R., The mechanical properties of muscle, «British Medical Bulletin», 1956, v. 12.
Скорость сокращения мышцы описывается уравнением
Эффективность двигателя или автомашины рассчитывают как процент потребляемой энергии, которая превращается в работу вместо тепла. В мышцах количество энергии, способной превращаться в работу, даже при наилучших условиях составляет менее 25% всей энергии, доставляемой к мышце (химической энергии питательных веществ), а остальная энергия превращается в тепло. Причина этой низкой эффективности связана с тем, что примерно половина энергии питательных веществ теряется во время образования АТФ, и только 40-45% энергии самой АТФ может позднее превратиться в работу.
Максимальная эффективность реализуется лишь при условии сокращения мышцы с умеренной скоростью. При медленном сокращении мышцы или без какого-либо ее укорочения во время сокращения освобождается небольшое количество поддерживающего тепла, хотя работа практически не выполняется, что снижает эффективность преобразования до нуля. Напротив, если сокращение слишком быстрое, большая доля энергии используется на преодоление вязкого трения внутри самой мышцы, и это также снижает эффективность сокращения. Обычно максимальная эффективность развивается, когда скорость сокращения составляет около 30%.
Характеристики сокращения целой мышцы
Многие особенности сокращения мышцы можно продемонстрировать на примере одиночных мышечных сокращений. Такие сокращения вызывают с помощью одиночного электрического возбуждения, иннервирующего мышцу нерва, или короткого электрического раздражения самой мышцы, что ведет к развитию одиночного сокращения, продолжающегося долю секунды.
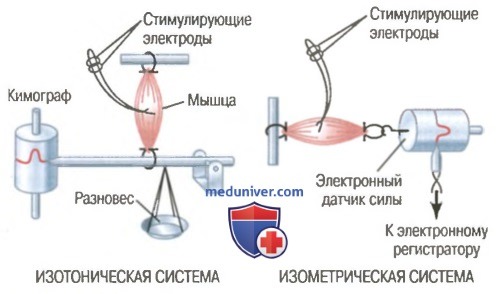 Изотоническая и изометрическая системы для регистрации мышечного сокращения.
Изотоническая и изометрическая системы для регистрации мышечного сокращения. 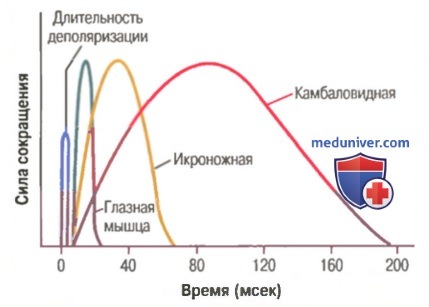 Длительность изометрических сокращений различных типов скелетных мышц млекопитающих. Показан также латентный период между потенциалом действия (деполяризацией) и мышечным сокращением.
Длительность изометрических сокращений различных типов скелетных мышц млекопитающих. Показан также латентный период между потенциалом действия (деполяризацией) и мышечным сокращением.
Изометрическое и изотоническое сокращение. Мышечное сокращение называют изометрическим, если мышца не укорачивается во время сокращения, и изотоническим — если мышца укорачивается, но ее напряжение на протяжении всего сокращения остается постоянным.
В изометрической системе мышца сокращается без уменьшения своей длины, а в изотонической системе мышца укорачивается против фиксированной нагрузки: мышца поднимает чашу весов с разновесом. Изометрическая система строго регистрирует изменения силы самого мышечного сокращения, а параметры изотонического сокращения зависят от нагрузки, против которой мышца сокращается, а также от инерции нагрузки. В связи с этим при сравнении функциональных особенностей различных типов мышц чаще всего используют изометрическую систему.
Особенности одиночных изометрических сокращений, зарегистрированных от разных мышц. В теле человека имеются много мышц разного размера — от очень маленькой стременной мышцы в среднем ухе, длиной в несколько миллиметров и диаметром около 1 мм, до очень большой четырехглавой мышцы, в 500000 раз крупнее стременной. При этом диаметр волокон может быть маленьким (10 мкм) или большим (80 мкм). Наконец, энергетика мышечных сокращений значительно варьирует от одной мышцы к другой. Поэтому не удивительно, что механические характеристики сокращений разных мышц различаются.
На рисунке показаны кривые регистрации изометрических сокращений трех типов скелетных мышц: глазной мышцы (длительность изометрического сокращения менее 1/40 сек), икроножной мышцы (длительность сокращения около 1/15 сек) и камбаловиднй мышцы (длительность сокращения примерно 1/3 сек). Интересно, что эти длительности сокращений приспособлены к функциям соответствующих мышц. Движения глаз должны быть чрезвычайно быстрыми, чтобы поддерживать фиксацию глаз на объекте для обеспечения ясного видения. Икроножная мышца должна сокращаться умеренно быстро, чтобы обеспечить скорость движения нижней конечности, достаточную для бега или прыжков. А камбаловидная мышца имеет дело в основном с медленными сокращениями для непрерывной длительной поддержки тела против силы тяжести.
Быстрые и медленные мышечные волокна. Как обсуждается в предыдущих статьях, посвященных спортивной физиологии, каждая мышца тела состоит из совокупности так называемых быстрых и медленных мышечных волокон, а также других волокон с переходными свойствами. В состав быстрореагирующих мышц входят в основном быстрые волокна и лишь небольшое число медленных. И наоборот, медленнореагирующие мышцы составлены главным образом из медленных волокон. Различия между этими двумя типами волокон следующие.
Быстрые волокна: (1) крупные волокна, обеспечивающие большую силу сокращения; (2) имеют хорошо развитый саркоплазматический ретикулум для быстрого выделения ионов кальция, инициирующих сокращение; (3) содержат большое количество гликолитических ферментов для быстрого освобождения энергии путем гликолиза; (4) имеют сравнительно бедное кровоснабжение, поскольку окислительный метаболизм имеет второстепенное значение; (5) содержат немного митохондрий также в связи со второстепенностью окислительного метаболизма.
Медленные волокна: (1) более мелкие волокна; (2) иннервируются также более мелкими нервными волокнами; (3) имеют хорошо развитую систему кровеносных сосудов и капилляров для доставки большого количества кислорода; (4) содержат значительно больше митохондрий для обеспечения высоких уровней окислительного метаболизма; (5) содержат большое количество миоглобина — железосодержащего белка, подобного гемоглобину эритроцитов. Миоглобин связывается с кислородом и хранит его до момента, когда в нем возникнет потребность (это также значительно увеличивает скорость транспорта кислорода в митохондрии). Миоглобин придает медленным волокнам красноватый вид, поэтому их называют красными волокнами, а из-за дефицита красного миоглобина в быстрых волокнах их называют белыми волокнами.
Видео физиология мышц и мышечного сокращения — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
http://slovaronline.com/browse/2469a0d0-7914-3f3f-a581-8f1836db0eb2/%D1%85%D0%B8%D0%BB%D0%BB%D0%B0-%D1%83%D1%80%D0%B0%D0%B2%D0%BD%D0%B5%D0%BD%D0%B8%D0%B5
http://meduniver.com/Medical/Physiology/520.html