Процесс клеточное дыхание его этапы кратко (Таблица)
Клеточное дыхание — это окислительный, с участием кислорода, распад органических питательных веществ, сопровождающийся образованием химически активных метаболитов и освобождением энергии, которые используются клетками для процессов жизнедеятельности.
Общее уравнение процесса дыхания имеет следующий вид:
где Q = 2878 кДж/моль.
Схема процесс клеточное дыхание
Дыхание — процесс многоступенчатый, в нем выделяют две основные стадии: гликолиз и кислородный этап (состоит из 3х подэтапов).

Таблица клеточное дыхание этапы
ATP (АТФ) — это аденозинтрифосфорная кислота, универсальный источник и переносчик энергии
NAD (НАД) — никотинамидадениндинуклеотидфосфата, кофермент
Ацетил-КоА — сложное органическое вещество ацетил-коэнзим А (СН3СО—S)
Пируват — это соли пировиноградной кислоты
Этапы клеточного дыхания
Первый этап: процесс гликолиза
Процесс гликолиза сложный и состоит примерно из десяти этапов. Глюкоза расщепляется («лизируется») на две молекулы пирувата. При этом образуются две молекулы АТР и две молекулы восстановленного кофермента. Эта стадия может протекать анаэробно, в анаэробных условиях (без кислорода или его недостатке) в результате гликолиза образуется молочная кислота (лактат), его еще называют брожение.
Второй этап: превращение пирувата в ацетил-КоА
Превращение пирувата в ацетил-КоА под действием пируватдегидрогеназного комплекса и направляет молекулу пирувата в цикл Кребса. Образуются две молекулы восстановленного кофермента. У эукариот процесс протекает в матриксе митохондрий.
Третий этап: цикл Кребса (цикл трикарбоновых кислот)
Цикл Кребса (трикарбоновых кислот или лимонной кислоты) представляет собой серию окислительных реакций. На каждом витке цикла образуется одна молекула АТР и четыре молекулы восстановленного кофермента. (На каждую молекулу глюкозы приходится два «оборота» цикла.) Это аэробная стадия.
Ацетил-КоА + 3NAD + + PAD + GDP + Фн + 2H2O + КоА-SH = 2КоА-SH + 3NADH + 3H + + PADН2 + GTP + 2CO2 (общее суммарное уравнение цикла)
Четвертый этап: окислительное фосфорилирование
Основное количество молекул АТP вырабатывается на этом этапе. Генерируется градиент протонов и его электрохимический потенциал используется для синтеза 32 молекул АТР. Аэробная стадия.
Кислород — это конечный акцептор восстановительного потенциала, возникающего при окислении органических молекул.
_______________
Источник информации:
1. Биология человека в диаграммах / В.Р. Пикеринг — 2003.
2. Биология для поступающих в вузы / Г.Л. Билич, В.А. Крыжановский. — 2008.
Суммарное уравнение дыхания два этапа
Процесс внешнего дыхания обусловлен изменением объема воздуха в легких в течение фаз вдоха и выдоха дыхательного цикла. При спокойном дыхании соотношение длительности вдоха к выдоху в дыхательном цикле равняется в среднем 1:1,3. Внешнее дыхание человека характеризуется частотой и глубиной дыхательных движений. Частота дыхания человека измеряется количеством дыхательных циклов в течение 1 мин и ее величина в покое у взрослого человека варьирует от 12 до 20 в 1 мин. Этот показатель внешнего дыхания возрастает при физической работе, повышении температуры окружающей среды, а также изменяется с возрастом. Например, у новорожденных частота дыхания равна 60—70 в 1 мин, а у людей в возрасте 25—30 лет — в среднем 16 в 1 мин. Глубина дыхания определяется по объему вдыхаемого и выдыхаемого воздуха в течение одного дыхательного цикла. Произведение частоты дыхательных движений на их глубину характеризует основную величину внешнего дыхания — вентиляцию легких. Количественной мерой вентиляции легких является минутный объем дыхания — это объем воздуха, который человек вдыхает и выдыхает за 1 мин. Величина минутного объема дыхания человека в покое варьирует в пределах 6—8 л. При физической работе у человека минутный объем дыхания может возрастать в 7—10 раз.
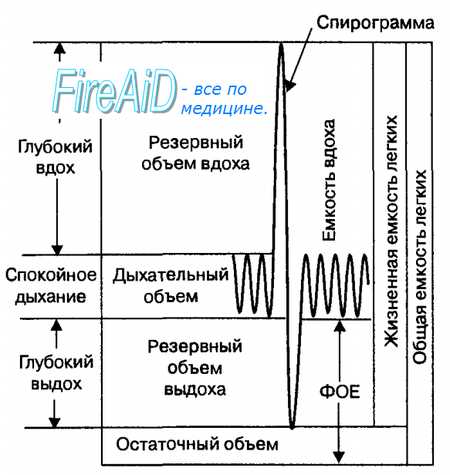 Рис. 10.5. Объемы и емкости воздуха в легких человека и кривая (спирограмма) изменения объема воздуха в легких при спокойном дыхании, глубоком вдохе и выдохе. ФОЕ — функциональная остаточная емкость.
Рис. 10.5. Объемы и емкости воздуха в легких человека и кривая (спирограмма) изменения объема воздуха в легких при спокойном дыхании, глубоком вдохе и выдохе. ФОЕ — функциональная остаточная емкость.
Легочные объемы воздуха. В физиологии дыхания принята единая номенклатура легочных объемов у человека, которые заполняют легкие при спокойном и глубоком дыхании в фазу вдоха и выдоха дыхательного цикла (рис. 10.5). Легочный объем, который вдыхается или выдыхается человеком при спокойном дыхании, называется дыхательным объемом. Его величина при спокойном дыхании составляет в среднем 500 мл. Максимальное количество воздуха, которое может вдохнуть человек сверх дыхательного объема, называется резервным объемом вдоха (в среднем 3000 мл). Максимальное количество воздуха, которое может выдохнуть человек после спокойного выдоха, называется резервным объемом выдоха (в среднем 1100 мл). Наконец, количество воздуха, которое остается в легких после максимального выдоха, называется остаточным объемом, его величина равна примерно 1200 мл.
Сумма величин двух легочных объемов и более называется легочной емкостью. Объем воздуха в легких человека характеризуется инспираторной емкостью легких, жизненной емкостью легких и функциональной остаточной емкостью легких. Инспираторная емкость легких (3500 мл) представляет собой сумму дыхательного объема и резервного объема вдоха. Жизненная емкость легких (4600 мл) включает в себя дыхательный объем и резервные объемы вдоха и выдоха. Функциональная остаточная емкость легких (1600 мл) представляет собой сумму резервного объема выдоха и остаточного объема легких. Сумма жизненной емкости легких и остаточного объема называется общей емкостью легких, величина которой у человека в среднем равна 5700 мл.
При вдохе легкие человека за счет сокращения диафрагмы и наружных межреберных мышц начинают увеличивать свой объем с уровня функциональной остаточной емкости, и его величина при спокойном дыхании составляет дыхательный объем, а при глубоком дыхании — достигает различных величин резервного объема вдоха. При выдохе объем легких вновь возвращается к исходному уровню функциональной остаточной емкости пассивно, за счет эластической тяги легких. Если в объем выдыхаемого воздуха начинает входит воздух функциональной остаточной емкости, что имеет место при глубоком дыхании, а также при кашле или чиханье, то выдох осуществляться за счет сокращения мышц брюшной стенки. В этом случае величина внутриплеврального давления, как правило, становится выше атмосферного давления, что обусловливает наибольшую скорость потока воздуха в дыхательных путях.
Аэробное и анаэробное дыхание растений
1. Аэробное дыхание
1.1 Окислительное фосфолирование
2. Анаэробное дыхание
2.1 Типы анаэробного дыхания
Дыхание присуще всем живым организмам. Оно представляет собой окислительный распад органических веществ, синтезированных в процессе фотосинтеза, протекающих с потреблением кислорода и выделением диоксида углерода. А.С. Фаминцын рассматривал фотосинтез и дыхание как две последовательные фазы питания растений: фотосинтез готовит углеводы, дыхание перерабатывает их в структурную биомассу растения, образуя в процессе ступенчатого окисления реакционноспособные вещества и освобождая энергию, необходимую для их превращения и процессов жизнедеятельности в целом. Суммарное уравнение дыхания имеет вид:
C  H
H  O + 6O
O + 6O  → 6CO + 6H O + 2875кДж.
→ 6CO + 6H O + 2875кДж.
Из этого уравнения становится ясно, почему именно скорость газообмена используют для оценки интенсивности дыхания. Оно было предложено в 1912 г. В. И. Палладиным, который считал, что дыхание состоит из двух фаз – анаэробной и аэробной. На анаэробном этапе дыхания, идущем в отсутствие кислорода, глюкоза окисляется за счет отнятия водорода (дегидрирования), который, по мнению ученого, передается на дыхательный фермент. Последний при этом восстанавливается. На аэробном этапе происходит регенерация дыхательного фермента в окислительную форму. В. И. Палладин впервые показал, что окисление сахара идет за счет непосредственного окисления его кислородом воздуха, поскольку кислород не встречается с углеродом дыхательного субстрата, а связано с его дегидрированием.
Существенный вклад в изучение сути окислительных процессов и химизма процесса дыхания внесли как отечественные (И.П. Бородин, А.Н.Бах, С.П. Костычев, В.И. Палладин), так и зарубежные (А.Л. Лавуазье, Г. Виланд, Г. Кребс) исследователи.
Жизнь любого организма неразрывно связана с непрерывным использованием свободной энергии, генерируемой при дыхании. Неудивительно, что изучению роли дыхания в жизни растения в последнее время отводят центральное место в физиологии растений.
1. Аэробное дыхание
Аэробное дыхание – это окислительный процесс, в ходе которого расходуется кислород. При дыхании субстрат без остатка расщепляется до бедных энергией неорганических веществ с высоким выходом энергии. Важнейшими субстратами для дыхания служат углеводы. Кроме того, при дыхании могут расходоваться жиры и белки.
Аэробное дыхание включает два основных этапа:
— бескислородный, в процессе, которого происходит постепенное расщепление субстрата с высвобождением атомов водорода и связыванием с коферментами (переносчиками типа НАД и ФАД);
— кислородный, в ходе которого происходит дальнейшее отщепление атомов водорода от производных дыхательного субстрата и постепенное окисление атомов водорода в результате переноса их электронов на кислород.
На первом этапе вначале высокомолекулярные органические вещества (полисахариды, липиды, белки, нуклеиновые кислоты и др.) под действием ферментов расщепляются на более простые соединения (глюкозу, высшие карбоновые кислоты, глицерол, аминокислоты, нуклеотиды и т.п.) Этот процесс происходит в цитоплазме клеток и сопровождается выделением небольшого количества энергии, которая рассеивается в виде тепла. Далее происходит ферментативное расщепление простых органических соединений.
Примером такого процесса является гликолиз – многоступенчатое бескислородное расщепление глюкозы. В реакциях гликолиза шестиуглеродная молекула глюкозы (С  ) расщепляется на две трехуглеродные молекулы пировиноградной кислоты (С
) расщепляется на две трехуглеродные молекулы пировиноградной кислоты (С  ). При этом образуется две молекулы АТФ, и выделяются атомы водорода. Последние присоединяются к переносчику НАД
). При этом образуется две молекулы АТФ, и выделяются атомы водорода. Последние присоединяются к переносчику НАД  (никотинамидадениндинклеотид), который переходит в свою восстановительную форму НАД ∙ Н + Н . НАД кофермент, близкий по своей структуре к НАДФ. Оба они представляют собой производные никотиновой кислоты – одного из витаминов группы В. Молекулы обоих коферментов электроположительны (у них отсутствует один электрон) и могут играть роль переносчика как электронов, так и атомов водорода. Когда акцептируется пара атомов водорода, один из атомов диссоциирует на протон и электрон:
(никотинамидадениндинклеотид), который переходит в свою восстановительную форму НАД ∙ Н + Н . НАД кофермент, близкий по своей структуре к НАДФ. Оба они представляют собой производные никотиновой кислоты – одного из витаминов группы В. Молекулы обоих коферментов электроположительны (у них отсутствует один электрон) и могут играть роль переносчика как электронов, так и атомов водорода. Когда акцептируется пара атомов водорода, один из атомов диссоциирует на протон и электрон:
Н → Н  + е
+ е  ,
,
а второй присоединяется к НАД или НАДФ целиком:
НАД + Н + [Н + е ] → НАД ∙ Н + Н.
Свободный протон позднее используется для обратного окисления кофермента. Суммарно реакция гликолиза имеет вид
C H O +2АДФ + 2Н  РО
РО  + 2 НАД →
+ 2 НАД →
2С Н О + 2АТФ + 2 НАД ∙ Н + Н + 2 HO
Продукт гликолиза – пировиноградная кислота (С Н О ) – заключает в себе значительную часть энергии, и дальнейшее ее высвобождение осуществляется в митохондриях. Здесь происходит полное окисление пировиноградной кислоты до CO и H O. Этот процесс можно разделить на три основные стадии:
1) окислительное декарбоксилирование пировиноградной кислоты;
2) цикл трикарбоновых кислот (цикл Кребса);
3) заключительная стадия окисления – электронтранспортная цепь.
На первой стадии пировиноградная кислота взаимодействует с веществом, которое называют коферментом А, в результате чего образуется ацетилкофермент а с высокоэнергетической связью. При этом от молекулы пировиноградной кислоты отщепляется молекула CO (первая) и атомы водорода, которые запасаются в форме НАД ∙ Н + Н .
Вторая стадия – цикл Кребса (рис. 1)

В цикл Кребса вступает ацетил–КоА, образованный на предыдущей стадии. Ацетил–КоА взаимодействует со щавелево-уксусной кислотой, в результате образуется шестиуглеродная лимонная кислота. Для этой реакции требуется энергия; ее поставляет высокоэнергетическая связь ацетил–КоА. В конце цикла щавелево-лимонная кислота регенерируется в прежнем виде. Теперь она способна вступить в реакцию с новой молекулой ацетил–КоА, и цикл повторяется. Суммарно реакция цикла может быть выражена следующим уравнением:
ацетил-КоА + 3H  O + 3НАД
O + 3НАД  + ФАД + АДФ + Н
+ ФАД + АДФ + Н  РО
РО  →
→
КоА + 2CO + 3НАД ∙ Н + Н +ФАД ∙ H + АТФ.
Таким образом, в результате распада одной молекулы пировиноградной кислоты в аэробной фазе (декарбоксилирование ПВК и цикла Кребса) выделяется 3CO , 4 НАД ∙ Н + Н , ФАД ∙ H . Суммарно реакцию гликолиза, окислительного декарбоксилирования и цикла Кребса можно записать в следующем виде:
C H  O + 6 H O + 10 НАД + 2ФАД →
O + 6 H O + 10 НАД + 2ФАД →
6CO + 4АТФ + 10 НАД ∙ Н + Н + 2ФАД ∙ H .
Третья стадия – электротранспортная цепь.
Пары водородных атомов, отщепляемые от промежуточных продуктов в реакциях дегидрирования при гликолизе и в цикле Кребса, в конце концов, окисляются молекулярным кислородом до H O с одновременным фосфолированием АДФ в АТФ. Происходит это тогда, когда водород, отделившийся от НАД ∙ H и ФАД ∙ H , передается по цепи переносчиков, встроенных во внутреннюю мембрану митохондрий. Пары атомов водорода 2Н можно рассматривать как 2 Н + 2е  . Движущей силой транспорта атомов водорода в дыхательной цепи является разность потенциалов.
. Движущей силой транспорта атомов водорода в дыхательной цепи является разность потенциалов.
С помощью переносчиков ионы водорода Н переносятся с внутренней стороны мембраны на ее внешнюю сторону, иначе говоря, из матрикса митохондрии в межмембранное пространство (рис. 2).

При переносе пары электронов от над на кислород они пересекают мембрану три раза, и этот процесс сопровождается выделением на внешнюю сторону мембраны шести протонов. На заключительном этапе протоны переносятся на внутреннюю сторону мембраны и акцептируются кислородом:
½ O + 2е  → O .
→ O .
В результате такого переноса ионов Н на внешнюю сторону мембраны митохондрий в перимитохондриальном пространстве создается концентрация их, т.е. возникает электрохимический градиент протонов .
Когда протонный градиент достигает определенной величины, ионы водорода из Н -резервуара движутся по специальным каналам в мембране, и их запас энергии используется для синтеза АТФ. В матриксе они соединяются с заряженными частичками О  , и образуется вода: 2Н + О²ˉ → H
, и образуется вода: 2Н + О²ˉ → H  O.
O.
1.1 Окислительное фосфолирование
Процесс образования АТФ в результате переноса ионов Н через мембрану митохондрии получил название окислительного фосфолирования. Он осуществляется при участии фермента АТФ-синтетазы. Молекулы АТФ-синтетазы располагаются в виде сферических гранул на внутренней стороне внутренней мембраны митохондрий.
В результате расщепления двух молекул пировиноградной кислоты и переноса ионов водорода через мембрану по специальным каналам синтезируется в целом 36 молекул АТФ (2 молекулы в цикле Кребса и 34 молекулы в результате переноса ионов Н  через мембрану).
через мембрану).
Суммарное уравнение аэробного дыхания можно выразить следующим образом:
C  H O + O + 6H O + 38АДФ + 38Н РО
H O + O + 6H O + 38АДФ + 38Н РО →
→
6CO + 12H O + 38АТФ
Совершенно очевидно, что аэробное дыхание прекратится в отсутствии кислорода, поскольку именно кислород служит конечным акцептором водорода. Если клетки не получают достаточного количества кислорода, все переносчики водорода вскоре полностью насытятся и не смогут передавать его дальше. В результате основной источник энергии дл образования АТФ окажется блокированным.
аэробное дыхание окисление фотосинтез
2. Анаэробное дыхание
Анаэробное дыхание. Некоторые микроорганизмы способны использовать для окисления органических или неорганических веществ не молекулярный кислород, а другие окисленные соединения, например, соли азотной, серной и угольной кислот, превращающиеся при этом в более восстановленные соединения. Процессы идут в анаэробных условиях, и их называют анаэробным дыханием:
2HNO  + 12Н
+ 12Н  → N + 6H O + 2Н
→ N + 6H O + 2Н
H SO + 8Н → H S + 4HO
У микроорганизмов, осуществляющих такое дыхание, конечным акцептором электронов будет не кислород а неорганическое соединения – нитриты, сульфаты и карбонаты. Таким образом, различия между аэробным и анаэробным дыханием заключается в природе конечного акцептора электронов.
2.1 Типы анаэробного дыхания
Основные типы анаэробного дыхания приведены в таблице 1. есть также данные об использовании бактериями в качестве акцепторов электронов Mn  , хроматов, хинонов и др.
, хроматов, хинонов и др.
Таблица 1 Типы анаэробного дыхания у прокариот (по: М.В Гусев, Л.А. Минеева 1992, с изменениями)
Конечный акцептор электронов
Нитратное дыхание и нитрификация
NO  , NO
, NO 
NO , NO, N  O, N
O, N
Сульфатное и серное дыхание
SO  , S
, S 
H S
Fe 
Fe 
CO
СН , ацетат
Свойство организмов переносить электроны на нитраты, сульфаты и карбонаты обеспечивает в достаточной степени полное окисление органического или неорганического вещества без использования молекулярного кислорода и обуславливает возможность получения большого количества энергии, чем при брожении. При анаэробном дыхании выход энергии только на 10% ниже. Чем при аэробном. Организмы, для которых характерно анаэробное дыхание, имеют набор ферментов электронтранспортной цепи. Но цитохромоксилаза в них заменяется нитратредуктазой (при использовании в качестве акцептора электронов нитрата) или аденилсульфатредуктазой (при использовании сульфата) или другими ферментами.
Организмы, способные осуществлять анаэробное дыхание за счет нитратов, — факультативные анаэробы. Организмы, использующие сульфаты в анаэробном дыхании, относятся к анаэробам.
Органические вещества из не органических зеленое растение образует только на свету. Эти вещества используются растением только для питания. Но растения не только питаются. Они дышат, как все живые существа. Дыхание происходит непрерывно днем о ночью. Дышат все органы растения. Растения дышат кислородом, а выделяют углекислый газ, как животные и человек.
Дыхание растений может происходить, как в темноте, так и на свету. Значит, на свету в растении протекают два противоположных процесса. Один процесс — фотосинтез, другой – дыхание. Во время фотосинтеза создаются органические вещества из неорганических и поглощается энергия солнечного света. Во время дыхания в растении расходуются органические вещества. А энергия, необходима для жизнедеятельности, освобождается. На свету в процессе фотосинтеза растения поглощают углекислый газ и выделяют кислород. Вместе с углекислым газом растения на свету поглощают из окружающего воздуха и кислород, необходимый растениям для дыхания, но в гораздо меньших количествах, чем выделяются при образовании сахара. Углекислого газа при фотосинтезе растения поглощают гораздо больше, чем выделяют его придыхании. Декоративные растения в комнате при хорошем освещении выделяют днем значительно больше кислорода, чем поглощают его в темноте ночью.
Дыхание во всех живых органов растения происходит непрерывно. Когда прекращается дыхание, растение, так же как и животное погибает.
1. Физиология и биохимия сельскохозяйственных растений Ф50/Н.Н. Третьяков, Е.И. Кошкин, Н.М. Макрушин и др.; под. ред. Н.Н. Третьякова. – М.; Колос, 2000 – 640 с.
2. Биология в экзаменационных вопросах и ответах Л44/ Лемеза Н.А., Камлюк Л.В.; 7-е изд. – М.: Айрис-пресс, 2003. – 512 с.
http://meduniver.com/Medical/Physiology/420.html
http://www.zinref.ru/000_uchebniki/00500biologia/000_lekcii_biologia_05/302.htm