Параграф 36. обмен глюкозо-6-фосфата
Автор текста – Анисимова Елена Сергеевна.
Авторские права защищены. Продавать текст нельзя.
Курсив не зубрить.
Замечания можно присылать по почте: exam_bch@mail.ru
https://vk.com/bch_5
ПАРАГРАФ 36. Сначала см. п. 30-35.
«Обмен ГЛЮКОЗО-6-ФОСФАТА».
В этом параграфе почти нет новых сведений, новые отмечены. В основном повтор того, чтобы было в параграфах с 30 по 35.
Метаболит глюкозо-6-фосфат есть в ряде процессов: в синтезе и распаде гликогена (п.31), в гликолизе (п.32) и ГНГ (п.33), в ПФП (п.35).
Как будет использоваться метаболит – зависит от того, какие ферменты могут на него подействовать, активны ли эти ферменты.
36. 1. Пути образования глюкозо-6-фосфата:
1 – из ГЛЮКОЗЫ при присоединении фосфата (что называется фосфорилированием и катализируется гексокиназой) – во всех клетках;
далее – или гликолиз (во всех клетках, п.32), или синтез гликогена (в печени и мышцах, п.31), или ПФП (п.35).
2 – из ГЛИКОГЕНА при гликогенолизе при стрессе и голоде – п.31,
3 – при ГЛЮКОНЕОГЕНЕЗЕ при стрессе и голоде – п.33; при этом оксалоацетат образуется из аминокислот благодаря реакциям ЦТК и переаминирования – см. п. 33 и 64 и 65.
4 – из фруктозы при их унификации – п.30.
Из галактозы можно получить глюкозо-6-фосфата, но при самой унификации галактозы нет этого метаболита, при унификации образуется глюкоза, а из неё можно получить глюкозо-6-фосфат.
36.2. Пути метаболизма (использования) глюкозо-6-фофата:
1 – ПРЕВРАЩЕНИЕ В ГЛЮКОЗУ при отщеплении фосфата (это называется дефосфорилированием и катализируется фосфатазой глюкозо-6-фосфата = глюкозо-6-фосфатазой);
реакция есть в ГНГ и гликогенолизе в печени (в мышцах нет этой реакции, потому что нет этого фермента),
протекает (как и ГНГ и гликогенолиз) ПРИ ГОЛОДЕ И СТРЕССЕ, чтобы образовать глюкозу, которая поступит в кровь для питания тканей и поддержания нормального уровня глюкозы в крови (гликемии),
2 – использование при ГЛИКОЛИЗЕ с последующим образованием лактата и 2 АТФ (если условия анаэробные) или СО2 и 32 АТФ.
3 – использование для синтеза ГЛИКОГЕНА (под действием инсулина – значит, при покое и сытости.
4 – использование при ПФП (под действием инсулина – значит, при покое и сытости);
при этом образованный при ПФП НАДФН обычно используется редуктазами и гидроксилазами,
но (это раньше не говорилось – новая информация) при сильном дефиците АТФ в клетке (что бывает при сильном стрессе, при гипоксии) НАДФН отдаёт водород не для этих реакций, а коферменту НАД+, превращая его в кофермент НАДН, который передаёт водород в дыхательную цепь для получения 2,5 АТФ.
Фермент, катализирующий передачу водорода от НАДФН на НАД+, называется трансгидрогеназой («переносчиком водорода, гидрогениума),
5 – использование для синтеза олигосахаридов и полисахаридов – п.38 и 39.
Глюкозо-6-фосфат→ фруктозо-6-фосфат
ГЛИКОЛИТИЧЕСКИЙ ПУТЬ ДЫХАТЕЛЬНОГО ОБМЕНА
Данный путь дыхательного обмена является наиболее распространенным и, в свою очередь, состоит из двух фаз. Первая фаза — анаэробная (гликолиз), вторая фаза — аэробная. Эти фазы локализованы в различныхкомпартментах клетки. Анаэробная фаза гликолиз — в цитоплазме, аэробная фаза — в митохондриях.
Анаэробная фаза дыхания (гликолиз)
В процессе гликолиза происходит преобразование молекулы гексозы до двух молекул пировиноградной кислоты (ПВК). Этот окислительный процесс может протекать в анаэробных условиях (в отсутствие кислорода) и идет через ряд этапов.
Прежде всего, для того чтобы подвергнуться дыхательному распаду, глюкоза должна быть активирована. Активация глюкозы происходит путем фосфорилирования шестого углеродного атома за счет взаимодействия с АТФ. Реакция идет в присутствии ионов магния и фермента гексокиназы:
глюкоза + АТФ→глюкозо-6-фосфат + АДФ.
Затем глюкозо-6-фосфат изомеризуется до фруктозо-6-фосфата. Процесс катализируется ферментом фосфоглюкоизомеразой:
глюкозо-6-фосфат→ фруктозо-6-фосфат.
Далее происходит еще одно фосфорилирование при участии АТФ. Фосфорная кислота присоединяется к первому углеродному атому молекулы фруктозы, процесс катализируется ферментом фосфофруктокиназой:
фруктозо-6-фосфат + АТФ→ фруктозо-1,6-дифосфат +АДФ.
Дальнейшие реакции, составляющие процесс гликолиза, складываются следующим образом: фруктозо-1,6-дифосфат расщепляется с образованием двух триоз, реакция катализируется ферментом альдолазой, которая состоит из четырех субъединиц и содержит свободные SH-группы. Реакция протекает по уравнению:
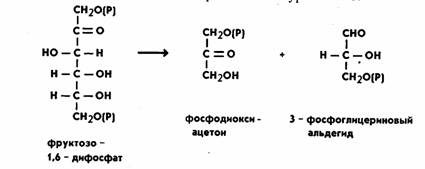
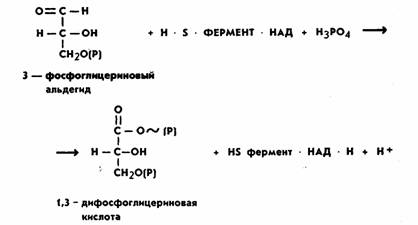
Молекула фосфодиоксиацетона при участии фермента триозофосфатизомеразы превращается также в 3-фосфоглицериновый альдегид (ФГА). Дальнейшим превращениям подвергается именно ФГА, окисляясь до 1,3-дифосфоглицериновой кислоты (ДФГК). Это важнейший этап гликолиза. Процесс идет с участием неорганического фосфата (H3РО4) и фермента глицеральдегид-3-фосфатдегидрогеназы.
Молекула этого фермента состоит из четырех идентичных субъединиц. Каждая субъединица представляет одиночную полипептидную цепь приблизительно из 220 аминокислотных остатков. Фермент содержит SH-группы и кофермент НАД, который взаимосвязан с ферментом на всем протяжении процесса. Сущность процесса заключается в окислении альдегидной группы ФГА в карбоксильную ДФГК. Окисление идет с выделением энергии. За счет энергии окисления при участии неорганического фосфата (Н3РО4) в молекуле ДФГК образуется макроэргическая фосфатная связь. Одновременно происходит восстановление кофермента НАД.
В целом реакция выглядит следующим образом:
На следующем этапе за счет имеющейся макроэргической связи в 1,3-дифосфоглицериновой кислоте образуется АТФ. Процесс катализируется ферментом фосфоглицераткиназой:
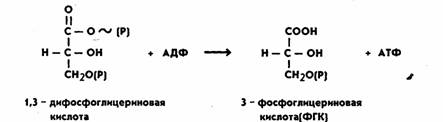
Таким образом, на этом этапе энергия окисления аккумулируется в форме энергии фосфатной связи АТФ. Затем 3-ФГК превращается в 2-ФГК, иначе говоря, фосфатная группа переносится из положения 3 в положение 2. Реакция катализируется ферментом фосфоглицеромутазой и идет в присутствии магния:
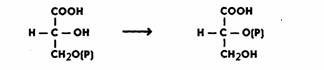
Далее происходит дегидратация ФГК. Реакция идет при участии фермента енолазы в присутствии ионов Mg2+ или Мn2+. Дегидратация сопровождается перераспределением энергии внутри молекулы, в результате чего возникает макроэргическая связь. Образуется фосфоенолпировиноградная кислота (ФЕП):
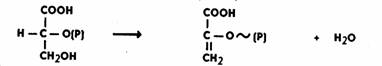
Затем фермент пируваткиназа переносит богатую энергией фосфатную группу на АДФ с образованием АТФ и пировиноградной кислоты. Для протекания реакции необходимо присутствие ионов Mg2+ или Мn2+:
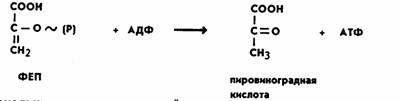
Поскольку при распаде одной молекулы глюкозы образуются две молекулы ФГА, то все реакции повторяются дважды. Таким образом, суммарное уравнение гликолиза следующее:
глюкоза + 2АТФ+ 2НАД+ + 2Фн + 4 АДФ → 2 пирувата + 4 АТФ + 2 НАД·Н2 + 2АДФ.
В результате процесса гликолиза образуются четыре молекулы АТФ, однако две из них покрывают расход на первоначальное активирование субстрата. Следовательно, накапливаются две молекулы АТФ. Образование АТФ в процессе гликолиза носит название субстратного фосфорилирования, поскольку макроэргические связи возникают на молекуле окисляемого субстрата. Если считать, что при распаде АТФ на АДФ и Фн выделяется 30,6 кДж, то за период гликолиза накапливается в макроэргических фосфатных связях всего 61,2 кДж. Прямые определения показывают, что распад молекулы глюкозы до пировиноградной кислоты сопровождается выделением 586,6 кДж. Следовательно, энергетическая эффективность гликолиза невелика. Кроме того, образуется 2НАД·Н2. НАД·Н2 вступает в дыхательную цепь, что приводит к дополнительному образованию АТФ. Образовавшиеся две молекулы пировиноградной кислоты вступают в аэробную фазу дыхания.
Синтез гликогена (гликогенез)
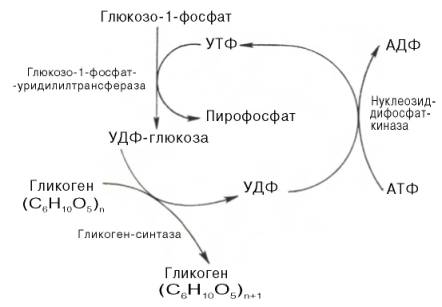
Прежде всего глюкоза подвергается фосфорилированию при участии фермента гексокиназы, а в печени – и глюкокиназы. Далее глюкозо-6-фосфат под влиянием фермента фосфоглюкомутазы переходит в глюкозо-1-фос-фат:
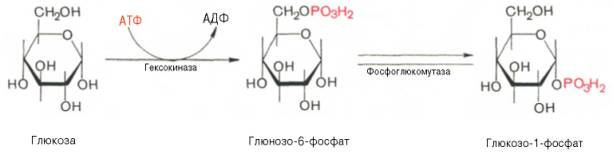
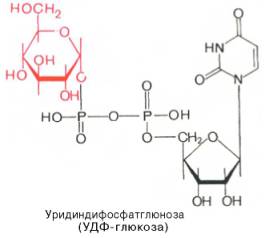
Образовавшийся глюкозо-1-фосфат уже непосредственно вовлекается в синтез гликогена. На первой стадии синтеза глюкозо-1-фосфат вступает во взаимодействие с УТФ (уридинтрифосфат), образуя уридиндифосфатглюкозу (УДФ-глюкоза) и пирофосфат. Данная реакция катализируется ферментом глюкозо-1-фосфат-уридилилтрансферазой (УДФГ-пирофосфорилаза):
Глюкозо-1-фосфат + УТФ УДФ-глюкоза + Пирофосфат.
На второй стадии – стадии образования гликогена – происходит перенос глюкозного остатка, входящего в состав УДФ-глюкозы, на глюкозидную цепь гликогена («затравочное» количество). При этом образуется α-(1–>4)-связь между первым атомом углерода добавляемого остатка глюкозы и 4-гидроксильной группой остатка глюкозы цепи. Эта реакция катализируется ферментом гликогенсинтазой. Необходимо еще раз подчеркнуть, что реакция, катализируемая гликогенсинтазой, возможна только при условии, что полисахаридная цепь уже содержит более 4 остатков D-глю-козы.
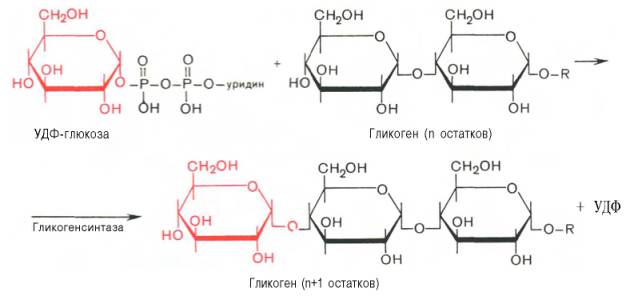
Образующийся УДФ затем вновь фосфорилируется в УТФ за счет АТФ, и таким образом весь цикл превращений глюкозо-1-фосфата начинается сначала.
В целом образование α-1,4-глюкозидной ветви («амилозной» ветви) гликогена можно представить в виде следующей схемы:
Установлено, что гликогенсинтаза неспособна катализировать образование α-(1–>6)-связи, имеющейся в точках ветвления гликогена. Этот процесс катализирует специальный фермент, получивший название гли-когенветвящего фермента, или амило-(1–>4)–>(1–>6)-трансглюкозидазы. Последний катализирует перенос концевого олигосахаридного фрагмента, состоящего из 6 или 7 остатков глюкозы, с нередуцирующего конца одной из боковых цепей, насчитывающей не менее 11 остатков, на 6-гидроксиль-ную группу остатка глюкозы той же или другой цепи гликогена. В результате образуется новая боковая цепь.
Ветвление повышает растворимость гликогена. Кроме того, благодаря ветвлению создается большое количество невосстанавливающих концевых остатков, которые являются местами действия гликогенфосфорилазы и гликогенсинтазы.
Таким образом, ветвление увеличивает скорость синтеза и расщепления гликогена.
Благодаря способности к отложению гликогена (главным образом в печени и мышцах и в меньшей степени в других органах и тканях) создаются условия для накопления в норме некоторого резерва углеводов. При повышении энерготрат в организме в результате возбуждения ЦНС обычно происходят усиление распада гликогена и образование глюкозы.
Помимо непосредственной передачи нервных импульсов к эффекторным органам и тканям, при возбуждении ЦНС повышаются функции ряда желез внутренней секреции (мозговое вещество надпочечников, щитовидная железа, гипофиз и др.), гормоны которых активируют распад гликогена, прежде всего в печени и мышцах (см. главу 8).
Как отмечалось, эффект катехоламинов в значительной мере опосредован действием цАМФ, который активирует протеинкиназы тканей. При участии последних происходит фосфорилирование ряда белков, в том числе гликогенсинтазы и фосфорилазы b – ферментов, участвующих в обмене углеводов. Фосфорилированный фермент гликогенсинтаза сам по себе малоактивен или полностью неактивен, но в значительной мере активируется положительным модулятором глюкозо-6-фосфатом, который увеличивает Vmaxфермента. Эта форма гликогенсинтазы называется D-формой, или зависимой (dependent) формой, поскольку ее активность зависит от глюкозо-6-фосфата. Дефосфорилированная форма гликоген-синтазы, называемая также I-формой, или независимой (independent) формой, активна и в отсутствие глюкозо-6-фосфата.
Таким образом, адреналин оказывает двойное действие на обмен углеводов: ингибирует синтез гликогена из УДФ-глюкозы, поскольку для проявления максимальной активности D-формы гликогенсинтазы нужны очень высокие концентрации глюкозо-6-фосфата, и ускоряет распад гликогена, так как способствует образованию активной фосфорилазы а. В целом суммарный результат действия адреналина состоит в ускорении превращения гликогена в глюкозу.
http://mydocx.ru/2-113019.html
http://xumuk.ru/biologhim/143.html