Потенциал покоя. Уравнение Гольдмана.
Между внутренней и внешней стороной клетки всегда есть разность электрических потенциалов. Эта разность в состоянии физиологического покоя клетки называется потенциалом покоя.
Установлено, что возникновение потенциала покоя обусловлено в основном наличием концентрационного градиента ионов калия. Эта точка зрения базируется на том, что ионы калия внутри клетки находятся в свободном состоянии, т.е. не связаны с другими ионами и молекулами и могут свободно диффундировать. Ионы калия диффундируют по концентрационному градиенту через клеточную мембрану и остаются на ее внутренней стороне. Т.к. ионы калия имеют «+» заряд, а анионы «-» заряд, то внешняя поверхность мембраны заряжается положительно, а внутренняя отрицательно.

Понятно, что диффузия продолжается только до тех пор, пока не установится равновесие между силами возникающего электрического поля и силами диффузии.
Мембрана в состоянии покоя проницаема не только для ионов калия, но и в небольшой степени для ионов натрия и хлора. Мембранный потенциал клеток представляет собой результирующую электродвижущих сил, генерируемых всеми этими каналами диффузии. Возникающий при этом потенциал описывается уравнением Гольдмана – Ходжкина – Катца:

где  – коэффициенты проницаемости мембраны для ионов
– коэффициенты проницаемости мембраны для ионов  .
.
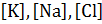 — концентрация ионов внутри (
— концентрация ионов внутри (  ) и вне (
) и вне (  ) клетки.
) клетки.
Таким образом, величина потенциала покоя клетки определяется двумя основными факторами:
1. соотношением концентраций проникающих через покоящуюся мембрану катионов и анионов;
2. соотношением проницаемостей мембраны для этих ионов.
Состояние покоя характеризуется следующим отношением ионных проницаемостей:
Уравнение Гольдмана переходит в уравнение Нернста, если мембрана проницаема для одного вида ионов. Следует отметить, что наличие обменной диффузии не влияет на мембранный потенциал.
Наиболее распространенный метод измерения биопотенциала покоя – микроэлектродный. При этом микроэлектроды вводятся: первый непосредственно в цитоплазму клетки, второй – во внеклеточную жидкость. Установлено, что цитоплазма в состоянии покоя клеток всегда имеет отрицательный потенциал по отношению к потенциалу межклеточной жидкости и колеблется в пределах от -100 милливольт до -50 милливольт.
6.3. Потенциал действия. Потенциал действия аксона нервной клетки.Уравнение Ходжкина – Хаксли.
Общее изменение разности потенциалов между клеткой и средой, происходящее при возбуждении клетки, называется потенциалом действия. В его основе лежат изменения ионной проницаемости мембраны. Потенциалы действия обеспечивают проведение возбуждения по нервным волокнам и инициируют процессы сокращения мышечных и секреции железистых клеток.
Представим график потенциала действия аксона нервной клетки. Потенциал как функция времени показан в фиксированной точке аксона.

φ1= -70 мВ потенциал покоя
φ2= +40 мВ мембранный потенциал при возбуждении
φ3 = /φ1/+ φ2 =110 мВ потенциал действия
φ4=-30 мВ запаздывающий потенциал.
Представим схему развития и распространения потенциала действия по аксону нервной клетки:
1. Состояние покоя.

Положительные и отрицательные заряды равномерно распределены вдоль аксона. Потенциал покояφ1= -70 мВ.
2. Приложен стимул.
 Происходит увеличение проницаемости мембраны для ионов натрия, примерно в 5000 раз. Поток ионов натрия из межклеточного пространства перекрывает отрицательный потенциал клетки и разрушает локальный положительный потенциал. Этот потенциал возникает в доли секунды и вырастает до φ2=40мВ и называется потенциалом действияφ3=110 мВ
Происходит увеличение проницаемости мембраны для ионов натрия, примерно в 5000 раз. Поток ионов натрия из межклеточного пространства перекрывает отрицательный потенциал клетки и разрушает локальный положительный потенциал. Этот потенциал возникает в доли секунды и вырастает до φ2=40мВ и называется потенциалом действияφ3=110 мВ
3. Распространение потенциала действия.
 Перемена мембранного потенциала называется деполяризацией. Эффект деполяризации, возникший в одной точке, индуцирует деполяризацию с обеих сторон от возбужденной точки.
Перемена мембранного потенциала называется деполяризацией. Эффект деполяризации, возникший в одной точке, индуцирует деполяризацию с обеих сторон от возбужденной точки.
Возникшее однажды значение потенциала действия распространяется по аксону до тех пор, пока не деполяризуется весь участок.
4. Полная деполяризация участка аксона.
 Почти сразу же после деполяризации мембрана приходит в исходное состояние с низкой проницаемости для ионов натрия.
Почти сразу же после деполяризации мембрана приходит в исходное состояние с низкой проницаемости для ионов натрия.
5. Начало реполяризации.
 Происходит резкое увеличение проницаемости мембраны для ионов калия. Ионы калия диффундируют из клетки, и это приводит к понижению потенциала клетки до отрицательных значений.
Происходит резкое увеличение проницаемости мембраны для ионов калия. Ионы калия диффундируют из клетки, и это приводит к понижению потенциала клетки до отрицательных значений.
«Ворота» для калия остаются открытыми в течение всего процесса реполяризации и, следовательно, потенциал падает ниже потенциала покоя и составляет минус 100 мВ. Клетка приходит в исходное состояние за 10 мсек.
Таким образом, формирование потенциала действия обусловлено двумя ионными потоками через мембрану: поток ионов натрия внутрь клетки приводит к перезарядке мембраны, а противоположно направленный поток ионов калия обуславливает восстановление исходного потенциала покоя.
Общее изменение мембранного потенциала при возбуждении складывается из потенциала покоя и мембранного потенциала при возбуждении  .Потенциал действия рассчитывается по уравнению Ходжкина – Хаксли.
.Потенциал действия рассчитывается по уравнению Ходжкина – Хаксли.

В состоянии возбуждения, отвечающим генерации потенциала действия, отношение ионных проницаемостей, следующее:

Потенциал действия функционирует по принципу «все или ничего» т.е. процесс либо происходит до конца, либо не начинается. Если стимул имеет величину, достаточную для запускания потенциала действия, то потенциал имеет одинаковую амплитуду и форму независимо от типа, величины и продолжительности действия стимула.
Скорость, с которой потенциал действия распространяется вдоль нервного волокна, зависит от диаметра волокна и от того покрыт ли аксон слоем миелина (структура из липопротеидов). У небольших волокон без миелина скорость может быть меньше 0,5 м/с, в то время как для больших волокон, покрытых миелином, скорость может быть 130 м/с. Ясно, что нервные импульсы не распространяются подобно электрическим сигналам. Распространение нервного импульса – это сложный электрохимический процесс, детали которого подробно рассматриваются при изучении таких дисциплин, как биохимия, физиология, нервные болезни.
Потенциал покоя. Уравнение Нернста. Уравнение Гольдмана-Ходжкина-Катца
Потенциал покоя (ПП, мембранный потенциал покоя). ПП называется разность потенциалов между внутренней и наружной сторонами клеточной мембраны, возникающей в состоянии покоя клетки. У живых клеток в покое между внутренним содержимым клетки и наружным раствором существует отрицательная разность потенциалов (ПП) порядка 60—90 мВ, которая локализована на поверхностной мембране. Внутренняя сторона мембраны заряжена электроотрицательно по отношению к наружной. ПП обусловлен избирательной проницаемостью покоящейся мембраны для ионов К + (Ю. Бернштейн, 1902, 1912. А. Ходжкин и Б. Катц, 1947).
Предположим, что клетку с мембраной, проницаемой только для ионов калия, поместили в электролит, где их концентрация меньше, чем внутри клетки. Сразу после соприкосновения мембраны с раствором ионы калия начнут выходить из клетки наружу, как выходит газ из надутого шара. Но каждый ион несет с собой положительный электрический заряд, и чем больше ионов калия покинет клетку, тем более электроотрицательным станет ее содержимое. Поэтому на каждый ион калия, выходящий из клетки, будет действовать электрическая сила, препятствующая его движению наружу. В конце концов, установится равновесие, при котором электрическая сила, действующая на ион калия в канале мембраны, будет равна силе, обусловленной различием концентраций ионов калия внутри и вне клетки. Очевидно, что в результате такого равновесия между внутренним и наружным растворами появится разность потенциалов. При этом, если за нуль потенциала принять потенциал внешнего раствора, то потенциал внутри клетки будет отрицательным. Эта разность потенциалов – самое простое из наблюдаемых биоэлектрических явлений – носит название «потенциал покоя» клетки. В первом приближении величину ПП можно рассчитать с помощью формулы Нернста:
 ,
,
где  — концентрация ионов калия внутри клетки,
— концентрация ионов калия внутри клетки,  — концентрация ионов калия снаружи клетки.
— концентрация ионов калия снаружи клетки.
Подставляя численные значения в формулу Нернста, получим значение ПП порядка -87 мВ, что весьма близко к экспериментально измеренному значению ПП. Дальнейшие исследования показали, что вклад в формирование ПП вносят не только ионы калия, но и ионы натрия и хлора.
Концентрация К + в протоплазме примерно в 50 раз выше, чем во внеклеточной жидкости, поэтому, диффундируя из клетки, ионы выносят на наружную сторону мембраны положительные заряды, при этом внутренняя сторона мембраны, практически не проницаемой для крупных органических анионов, приобретает отрицательный потенциал. Поскольку проницаемость мембраны в покое для Na + примерно в 100 раз ниже, чем для К + , диффузия натрия из внеклеточной жидкости (где он является основным катионом) в протоплазму мала и лишь незначительно снижает ПП, обусловленный ионами К + .
В скелетных мышечных волокнах в возникновении потенциала покоя важную роль играют также ионы Cl — , диффундирующие внутрь клетки. Следствием ПП является ток покоя, регистрируемый между поврежденным и интактным участками нерва или мышцы при приложении отводящих электродов. Мембраны нервных и мышечных клеток (волокон) способны изменять ионную проницаемость в ответ на сдвиги мембранного потенциала. При увеличении ПП (гиперполяризация мембраны) проницаемость поверхностных клеточных мембран для Na + и К + падает, а при уменьшении ПП (деполяризация) она возрастает, причём скорость изменений проницаемости для Na + значительно превышает скорость увеличения проницаемости мембраны для К + . Более точные значения для ПП вычисляются по формуле Гольдмана-Ходжкина-Катца (ГХК), которая учитывает проницаемость КМ в покое не только для ионов калия, но и для ионов натрия и хлора:
 ,
,
где PK, PNa, PCl – проницаемость КМ для ионов калия, натрия и хлора, выражения […]i и […]0 обозначают концентрации соответствующих молекул внутри и вне клетки.
В качестве примера вычислим величину ПП для гигантского аксона кальмара. Концентрации ионов внутри и вне аксона приведены ниже (см. таблицу).
| Ион | Концентрация (моль на 1 кг воды) |
| Внутри (i) | Вне (0) |
| Na + | |
| K + | |
| Cl — |
В состоянии покоя при физиологических условиях соотношение коэффициентов проницаемости равно:
Диффузия ионов калия и хлора идет через КМ в обе стороны. Натрий за счет механизма пассивного транспорта проникает внутрь аксона и за счет активного транспорта выносится из клетки. Следовательно, основной вклад в формирование ПП вносят ионы калия и хлора. Подставив численные значения проницаемостей и концентраций для ионов калия хлора в формулу Гольдмана-Ходжкина-Катца для температуры t=30 0 C вычислим:

Значение ПП, вычисленное по формуле Нернста, равно:

Формула Нернста дает несколько заниженное значение потенциала покоя, а формула ГХК приводит к более реалистичным значениям ПП, измеренным экспериментально на крупных клетках.
Следует также отметить, что ни формула Нернста, ни формула ГХК не учитывают механизма активного транспорта.
Формула Томаса для ПП учитывает работу электрогенных ионных Na-K- насосов и имеет вид:
 ,
,
где m – отношение количества ионов натрия к количеству ионов калия, переносимых натрий-калиевым насосом через КМ. Наиболее распространенный режим работы Na + -K + -АТФ-азы наблюдается при m=3/2. В уравнении Томаса отсутствуют члены PCl[Cl — ], так как нет активного транспорта для ионов хлора через КМ.
Численное значение ПП по формуле Томаса равно:
 .
.
Коэффициент m, применяемый в уравнении Томаса, усиливает вклад градиента концентрации ионов калия в формирование ПП. Поэтому ПП, рассчитанный по формуле Томаса, по абсолютному значению меньше ПП, рассчитанного по формуле ГХК. Значение ПП, полученное с помощью формулы Томаса, хорошо совпадает со значениями ПП, измеренными экспериментально на мелких клетках.
Механизм возникновения потенциала покоя на биологических мембранах. Формула Нернста, формула Гольдмана
Потенциал покоя – это стационарная разность потенциалов между внешней и внутренней средой клетки, существующая на наружной мембране клетки в невозбужденном состоянии.
Зарегистрировать эту трансмембранную разность потенциалов можно при использовании микроэлектродной техники. Стеклянный микроэлектрод представляет собой стеклянную микропипетку с оттянутым очень тонким кончиком (диаметр кончика 0,1-0,5 мкм), заполненную раствором электролита (обычно 3 М раствором KCl). Таким электродом можно проколоть наружную мембрану клетки, не повредив ее. Второй электрод (электрод сравнения) помещают в раствор у наружной поверхности клетки. Регистрирующее устройство, содержащее усилитель постоянного тока, позволяет измерить трансмембранную разность потенциалов. При этом в нервных и мышечных волокнах различных животных регистрируется разность потенциалов, равная примерно 80-90 мВ (за исключением гладкомышечных клеток, потенциал покоя которых ниже (- 30 мВ), причем внутренняя поверхность клеточной мембраны имеет отрицательный потенциал по отношению к внешней.
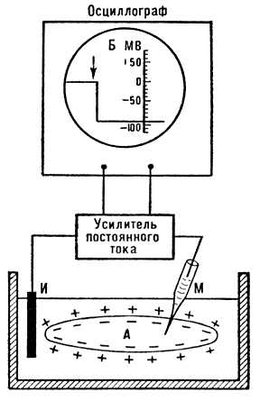
Как же реализуются обязательные условия биоэлектрогенеза на наружной мембране клетки в состоянии покоя?
1) В цитоплазме позвоночных животных преобладают калиевые соли высокомолекулярных соединений (кислот), тогда как в межклеточной среде гораздо выше концентрация натриевых солей неорганических кислот. Например, в гигантском аксоне кальмара (очень удобном для исследований биоэлектрогенеза в связи с большим диаметром) внутриклеточная концентрация К + равна 410 ммоль/л, во внеклеточной среде – 10 ммоль/л. Соответственно, концентрация натрия составляет 49 и 460 ммоль/л. Сходные соотношения наблюдаются и в других клетках.
2) В покое проницаемость наружной мембраны клетки для ионов калия значительно больше, чем для натрия, и больше, чем для ионов хлора:
Например, для аксона кальмара:
Эта проницаемость обусловлена наличием в наружной мембране так называемых потенциалнезависимых каналов, избирательно пропускающих ионы калия, натрия или хлора.
Если концентрация какого-либо иона внутри клетки отлична от концентрации этого иона снаружи и мембрана проницаема для этого иона, возникает поток заряженных частиц через мембрану, вследствие чего электрическая нейтральность системы нарушается, образуется разность потенциалов между наружной и внутренней средой, которая препятствует дальнейшему переносу ионов через мембрану. При установлении равновесия между концентрационным и электрическим градиентом выравниваются значения электрохимических потенциалов по разные стороны мембраны и устанавливается равновесный потенциал для этого иона, который рассчитывается по формуле Нернста.
 μ = μ0 + RTlnC + zFφ
μ = μ0 + RTlnC + zFφ
φ равн. = φi – φe = 
Если принять, что мембранный потенциал обусловлен только переносом ионов калия, то калиевый равновесный потенциал равен:
φ равн. =  -4 % ионов калия от общего их содержания внутри клетки (изменение концентрации калия всего на 2 . 10 -3 ммоль/л). Это пренебрежимое малое количество ионов по сравнению с общим их количеством в клетке.
-4 % ионов калия от общего их содержания внутри клетки (изменение концентрации калия всего на 2 . 10 -3 ммоль/л). Это пренебрежимое малое количество ионов по сравнению с общим их количеством в клетке.
Причина расхождения рассчитанного по формуле Нернста и экспериментального значения мембранного потенциала состоит в том, что не учтена проницаемость мембраны для других ионов (натрия и хлора). Более точно с экспериментальными данными совпадает результат расчета по уравнению Гольдмана:

В числителе выражения, представленного под знаком логарифма, представлены концентрации калия и натрия внутри клетки и концентрация хлора снаружи, в знаменателе – наоборот, это связано с тем, что ионы хлора заряжены отрицательно.
Мембранный потенциал, рассчитанный по уравнению Гольдмана, по абсолютной величине меньше мембранного потенциала, рассчитанного по уравнению Нернста, и ближе к экспериментальным значениям.
И формула Нернста, и формула Гольдмана не учитывают активного транспорта ионов через мембрану – наличия в мембране электрогенного биологического насоса – K + -Na + -АТФазы, перекачивающей калий внутрь клетки, а натрий наружу в неравновесных соотношениях. Чаще всего АТФаза работает в режиме 3Na:2K, то есть за счет работы АТФазы мембранный потенциал больше по абсолютной величине, чем мембранный потенциал, рассчитанный по уравнению Гольдмана.
Если прекратить поступление Na + в клетку, например, путем замещения внеклеточного натрия на такой неспособный к диффузии катион, как холин, то МП будет близок к калиевому равновесному потенциалу.
Нарушение работы K + -Na + -АТФазы приводит к уменьшению мембранного потенциала; в этом случае МП лучше описывается уравнением Гольдмана.
Повреждение клеточной мембраны приводит к повышению проницаемости мембраны для всех ионов. В этом случае разница в проницаемостях сглаживается, МП уменьшается.
Для сильно поврежденных клеток МП еще меньше, но отрицательный мембранный потенциал сохраняется за счет наличия в клетке полианионов – отрицательно заряженных белков, нуклеиновых кислот и других крупных молекул, которые не могут проникнуть через мембрану (доннановский потенциал).
http://megalektsii.ru/s23176t4.html
http://helpiks.org/5-71086.html