Уравнение хилла для мышечного сокращения
Эффективность двигателя или автомашины рассчитывают как процент потребляемой энергии, которая превращается в работу вместо тепла. В мышцах количество энергии, способной превращаться в работу, даже при наилучших условиях составляет менее 25% всей энергии, доставляемой к мышце (химической энергии питательных веществ), а остальная энергия превращается в тепло. Причина этой низкой эффективности связана с тем, что примерно половина энергии питательных веществ теряется во время образования АТФ, и только 40-45% энергии самой АТФ может позднее превратиться в работу.
Максимальная эффективность реализуется лишь при условии сокращения мышцы с умеренной скоростью. При медленном сокращении мышцы или без какого-либо ее укорочения во время сокращения освобождается небольшое количество поддерживающего тепла, хотя работа практически не выполняется, что снижает эффективность преобразования до нуля. Напротив, если сокращение слишком быстрое, большая доля энергии используется на преодоление вязкого трения внутри самой мышцы, и это также снижает эффективность сокращения. Обычно максимальная эффективность развивается, когда скорость сокращения составляет около 30%.
Характеристики сокращения целой мышцы
Многие особенности сокращения мышцы можно продемонстрировать на примере одиночных мышечных сокращений. Такие сокращения вызывают с помощью одиночного электрического возбуждения, иннервирующего мышцу нерва, или короткого электрического раздражения самой мышцы, что ведет к развитию одиночного сокращения, продолжающегося долю секунды.
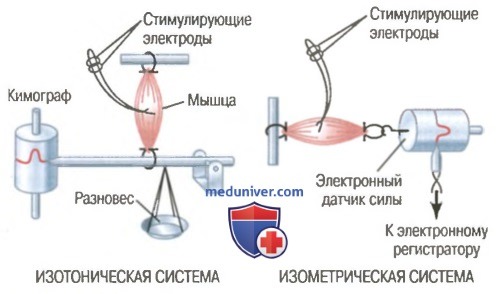 Изотоническая и изометрическая системы для регистрации мышечного сокращения.
Изотоническая и изометрическая системы для регистрации мышечного сокращения. 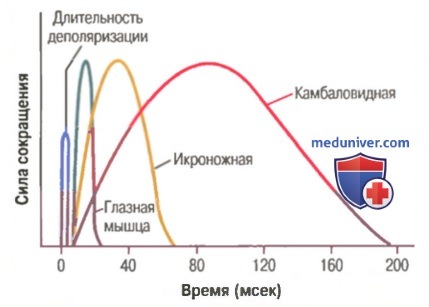 Длительность изометрических сокращений различных типов скелетных мышц млекопитающих. Показан также латентный период между потенциалом действия (деполяризацией) и мышечным сокращением.
Длительность изометрических сокращений различных типов скелетных мышц млекопитающих. Показан также латентный период между потенциалом действия (деполяризацией) и мышечным сокращением.
Изометрическое и изотоническое сокращение. Мышечное сокращение называют изометрическим, если мышца не укорачивается во время сокращения, и изотоническим — если мышца укорачивается, но ее напряжение на протяжении всего сокращения остается постоянным.
В изометрической системе мышца сокращается без уменьшения своей длины, а в изотонической системе мышца укорачивается против фиксированной нагрузки: мышца поднимает чашу весов с разновесом. Изометрическая система строго регистрирует изменения силы самого мышечного сокращения, а параметры изотонического сокращения зависят от нагрузки, против которой мышца сокращается, а также от инерции нагрузки. В связи с этим при сравнении функциональных особенностей различных типов мышц чаще всего используют изометрическую систему.
Особенности одиночных изометрических сокращений, зарегистрированных от разных мышц. В теле человека имеются много мышц разного размера — от очень маленькой стременной мышцы в среднем ухе, длиной в несколько миллиметров и диаметром около 1 мм, до очень большой четырехглавой мышцы, в 500000 раз крупнее стременной. При этом диаметр волокон может быть маленьким (10 мкм) или большим (80 мкм). Наконец, энергетика мышечных сокращений значительно варьирует от одной мышцы к другой. Поэтому не удивительно, что механические характеристики сокращений разных мышц различаются.
На рисунке показаны кривые регистрации изометрических сокращений трех типов скелетных мышц: глазной мышцы (длительность изометрического сокращения менее 1/40 сек), икроножной мышцы (длительность сокращения около 1/15 сек) и камбаловиднй мышцы (длительность сокращения примерно 1/3 сек). Интересно, что эти длительности сокращений приспособлены к функциям соответствующих мышц. Движения глаз должны быть чрезвычайно быстрыми, чтобы поддерживать фиксацию глаз на объекте для обеспечения ясного видения. Икроножная мышца должна сокращаться умеренно быстро, чтобы обеспечить скорость движения нижней конечности, достаточную для бега или прыжков. А камбаловидная мышца имеет дело в основном с медленными сокращениями для непрерывной длительной поддержки тела против силы тяжести.
Быстрые и медленные мышечные волокна. Как обсуждается в предыдущих статьях, посвященных спортивной физиологии, каждая мышца тела состоит из совокупности так называемых быстрых и медленных мышечных волокон, а также других волокон с переходными свойствами. В состав быстрореагирующих мышц входят в основном быстрые волокна и лишь небольшое число медленных. И наоборот, медленнореагирующие мышцы составлены главным образом из медленных волокон. Различия между этими двумя типами волокон следующие.
Быстрые волокна: (1) крупные волокна, обеспечивающие большую силу сокращения; (2) имеют хорошо развитый саркоплазматический ретикулум для быстрого выделения ионов кальция, инициирующих сокращение; (3) содержат большое количество гликолитических ферментов для быстрого освобождения энергии путем гликолиза; (4) имеют сравнительно бедное кровоснабжение, поскольку окислительный метаболизм имеет второстепенное значение; (5) содержат немного митохондрий также в связи со второстепенностью окислительного метаболизма.
Медленные волокна: (1) более мелкие волокна; (2) иннервируются также более мелкими нервными волокнами; (3) имеют хорошо развитую систему кровеносных сосудов и капилляров для доставки большого количества кислорода; (4) содержат значительно больше митохондрий для обеспечения высоких уровней окислительного метаболизма; (5) содержат большое количество миоглобина — железосодержащего белка, подобного гемоглобину эритроцитов. Миоглобин связывается с кислородом и хранит его до момента, когда в нем возникнет потребность (это также значительно увеличивает скорость транспорта кислорода в митохондрии). Миоглобин придает медленным волокнам красноватый вид, поэтому их называют красными волокнами, а из-за дефицита красного миоглобина в быстрых волокнах их называют белыми волокнами.
Видео физиология мышц и мышечного сокращения — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Уравнение Хилла. Мощность одиночного сокращения
ГОСУДАРСТВЕННЫЙ МЕДИЦИНСКИЙ УНИВЕРСИТЕТ г. СЕМЕЙ
Методическое пособие по теме:
Приборы для измерения функции внешнего дыхания. Устройство и принцип работы.
Регистрация и анализ данных функциональных исследований
Составители: Крылова Л.А.
БИОФИЗИКА МЫШЕЧНОГО СОКРАЩЕНИЯ.
Мышечная активность – это одно из общих свойств высоко организованных живых организмов. Вся жизнедеятелъность человека связана с мышечной активностью. Независимо от назначения, особенностей строения и способов регуляции принцип работы различных мышц организма одинаков.
Мышечная клетка отличается от других возбудимых клеток таким специфическим свойством, как сократимость, т.е. способность генерировать механическое напряжение и укорачиваться.Кроме того, мышцы являются генератором тепла, причем не только при мышечной работе, холодовой дрожи, но и в режиме термогенеза.
Мышечная активность о процессе жизнедеятельности обеспечивает работы отдельных органов и целых систем: работу опорно-двигательного аппарата, легких, сосудистую активностъ, работу желудочно-кишечного тракта, сократительную способность сердца. Нарушение работы мышц (например, определяющих функционирование легких, сердца) может приводить к патологиям, а ее прекращение — даже к летальному исходу.
Структура поперечно-полосатой мышцы. Модель скользящих нитей.
Мышечная ткань представляет собой совокупность мышечных клеток (волокон), внеклеточного вещества (коллаген, эластина и др.) и густой сети нервных волокон и кровеносных сосудов. Мышцы по строению делятся на:гладкие— мышцы кишечника, стенки сосудов, и поперечно-полосатые— скелетные, мышцы сердца. Независимо от строения все они имеют близкие механические свойства, одинаковый механизм активации и
близкий химический состав.

Справа – схема расположения актина и миозина на поперечном срезе.
В миофибриллах (Рис.1) различают: А -зону — темные полосы, которые в поляризованном свете дают двойное лучепреломление, т.е. обладают свойством анизотропии (отсюда и название: А -зона), I -зону — светлые полосы, не дающие двойного лучепреломления, то есть изотропные(отсюда название: I -зона). В области I-зоны проходит темная узкая полоса –
Z-диск. Промежуток между двумя Z-дисками называется саркомероми является элементарной сократительной единицей мышечной клетки.
Саркомер— это упорядоченная система толстых и тонких нитей, расположенных гексагопально в поперечном сечении. Толстая нить имеет толщину — 12 нм и длину — 1,5 мкм и состоит из белка миозина. Тонкая нить имеет диаметр 8 нм, длину 1 мкм и состоит из белка актина, прикрепленного одним концом к Z-диску.

Рис.2. Микроструктура саркомера.
Актиновая нить состоит из двух закрученных один вокруг другого мономеров актина толщиной по 5 нм (рис. 2). Эта структура похожа на две нитки бус, скрученные по 14 бусин в витке. В цепях актина регулярно примерно через 40 нм встроены молекулы тропонина, а сама цепь охватывает нить тропомиозина. При сокращении мышцы тонкие нити вдвигаются между толстыми нитями. Происходит относительное скольжениенитей без изменения их длины. Этот процесс обусловлен взаимодействием особых выступов миозина — поперечных мостиковс активными центрами, расположенными на актине. Мостики отходят от толстой нити периодично на расстоянии 14,5 нм друг от друга.
В расслабленном состоянии миофибрил молекулы тропомиазина блокируют прикрепление поперечных мостиков к актиновым цепям (Рис.3.а).
 Мостик разомкнут. Замыкание. Усиление.
Мостик разомкнут. Замыкание. Усиление.


Рис. 3. Процесс активации мостика и генерации усилия в саркомере.
Ионы Са активируют мостики и открывают участки их прикрепления к актину (рис. б).
В результате мостики миозина прикрепляются к актиновым нитям, расщепляются молекулы АТФ и изменяется конфирмация мостиков: их головки поворачиваются внутрь саркомера (Рис.3. в). Это приводит к генерации силы, скольжению актина относительно толстой нити миозина к центру саркомера, что вызывает укорочение мышцы. После окончания активации мостик размыкается и саркомер возвращается в исходное состояние. При укорочении объем саркомера практически не меняется, следовательно, он становится толще, что и подтверждается на снимках поперечного сечения мышц с помощью электронной микроскопии. Каждый цикл замыкание — размыкание сопровождается расщеплением одной молекулы АТФ. Таким образом, актин — миозиновый комплекс
является механохимическим преобразователем энергии АТФ. Рассмотренная структура и последовательность процессов называется моделью скользящих нитей.
Впервые скольжение нитей в саркомере было обнаружено английским ученым Х.Хаксли. Он же сформулировал модель скользящих нитей. Существенный вклад в разработку теории скользящих нитей внес В.И. Дещеревский,
Представленная структурная модель скользящих нитей надежно подтверждена экспериментально и является опытным фактом, на котором основаны все современные теории мышечного сокращения. Эксперементальные данные о микроструктуре мышц были изучены с помощью электронной микроскопии, рентгеноструктурного анализа и метода дифракции синхротронного излучения.
Основные положения модели скользящих нитей:
1. Длины нитей актина и миозина в ходе сокращения не меняются.
2. Изменение длины саркомера при сокращении — результат относительного продольного смещения нитей актина и миозина.
3. Поперечные мостики, отходящие от миозина, могут присоединяться к комплементарнымцентрам актина.
4. Мостики прикрепляются к актину не одновременно.
5. Замкнувшиеся мостики подвергаются структурному переходу, при котором они развивают усилие, после чего происходит их размыкание.
6. Сокращение и расслабление мышцы состоит в нарастании и последующем уменьшении числа мостиков, совершающих цикл замыкaние — размыкание.
7. Каждый цикл связан с гидролизом одной молекулы АТФ.
8. Акты замыкания-размыкания мостиков происходят не зависимо друг от друга.
На рис. 4. представлена зависимость максимального значения развиваемой силы от степени перекрытия актиновых и миозиновых нитей.

 Возможность саркомера сократиться и развить усилие в большой степени зависит от начальных условий. Если саркомер изначально растянут (его длина 3,65 мкм), то мостики не перекрываются актиновыми нитями и при стимуляции такого элемента усилие не формируется(стрелка 1 на рис. 4, а и фрагмент 1 на рис. 4, б). Если саркомер находится в рабочем начальном состоянии (размер саркомера 2,2 мкм), то при стимуляции он разовьет максимальную силу (стрелка 2, рис. 4, а и фрагмент 2 рис. 4, б). Если начальный размер саркомера слишком короток, генерация усилия уменьшается (стрелка и фрагмент 5).
Возможность саркомера сократиться и развить усилие в большой степени зависит от начальных условий. Если саркомер изначально растянут (его длина 3,65 мкм), то мостики не перекрываются актиновыми нитями и при стимуляции такого элемента усилие не формируется(стрелка 1 на рис. 4, а и фрагмент 1 на рис. 4, б). Если саркомер находится в рабочем начальном состоянии (размер саркомера 2,2 мкм), то при стимуляции он разовьет максимальную силу (стрелка 2, рис. 4, а и фрагмент 2 рис. 4, б). Если начальный размер саркомера слишком короток, генерация усилия уменьшается (стрелка и фрагмент 5).
Рис. 4. Зависимость максимального значения развиваемой силы Р, развиваемой при изометрическом сокращении от начальной длины саркомера l (а) и степени перекрытия актиновых и миозиновых нитей (б).
Биомеханика мышцы
Мышцы можно представить как сплошную среду, т.е. среду, состоящую из большого числа элементов, взаимодействующих между собой без соударений и находящихся в поле внешних сил. Мышца одновременно обладает свойством упругости и вязкости, то есть является вязко-упругой средой. Для такой среды используются законы классической механики.
Фундаментальными понятиями механики сплошных сред являются деформация, напряжение, упругость, вязкость, а также энергия и температура.
а). Упругость —свойство тел менять размеры и форму под действием сил и самопроизвольно восстанавливать их при прекращении внешних воздействий.
Упругость тел обусловлена силами взаимодействия его атомов и молекул. При снятии внешнего воздействия тело самопроизвольно возвращается в исходное состояние.
б) Вязкость – внутренне трение среды.
в). Вязкоупругость– это свойство материалов твердых тел сочетать упругость и вязкость.
г) Деформация относительное изменение длины:  ,
,
где l -начальная длина,  — значение удлинения, может изменять знак.
— значение удлинения, может изменять знак.
д) Напряжение механическое  — мера внутренних сил, возникающих при деформации материала. Для однородного стержня:
— мера внутренних сил, возникающих при деформации материала. Для однородного стержня:  , где S
, где S
площадь сечения, F — сила,  приложенная к стержню.
приложенная к стержню.
Упругая деформация возникает и исчезает одновременно с нагрузкой и не сопровождается рассеянием энергии. Для упругой деформации справедлив закон Гука: 
где Е — модуль Юнга, определяемый природой вещества. При растяжении различных материалов, в общем случае,  . При малых растяжениях считают Е=const.
. При малых растяжениях считают Е=const.
Таблица значений модуля Юнга для различных материалов.
| Материал | Модуль Юнга (Па) |
| Эластин |  |
| Коллаген |  |
| Кость |  |
| Резина |  |
| Дуб | 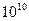 |
| Сталь |  |
Эластин — упругий белок позвоночных, находится, в основном, в стенках артерий. Коллаген — волокнистый белок. В мышцах 20 % всех белков – коллаген, он находится также а сухожилиях, хрящах, кости.
В случае вязкой среды напряжение  определяется скоростью деформации d
определяется скоростью деформации d  /dt,
/dt,  , где
, где  — коэффициент вязкости среды.
— коэффициент вязкости среды.
Для вязкоупругой деформации характерна явная зависимость от процесса нагружения во времени, причём при снятии нагрузки деформация с течением некоторого времени самопроизвольно стремится к нулю.
Пассивные механические свойства вязкоупругой среды можно моделировать сочетанием упругих и вязких элементов. Мышцане является ни чисто упругим, ни чисто вязким элементом. Мышца — вязкоупругий элемент.
Пассивное растяжение. На основании расчетных и экспериментальных данных показано, что наиболее простой моделью, дающей достаточно хорошее приближение кмеханическим свойствам мышцы, является трехкомпонентная модель Хилла (Рис. 5).
 Для установления характерной зависимости деформации мышцы во времени при приложении к ней мгновенного воздействия упростим модель (рис. 5). Допустим, что элемент Е2, отсутствует (
Для установления характерной зависимости деформации мышцы во времени при приложении к ней мгновенного воздействия упростим модель (рис. 5). Допустим, что элемент Е2, отсутствует (  ). Тогда для вязкоупругой среды напряжение в системе будет определиться упругой
). Тогда для вязкоупругой среды напряжение в системе будет определиться упругой  с и вязкой составляющими:
с и вязкой составляющими:  + ,
+ , 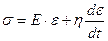
Найдем зависимость (t), решая дифференциальное уравнение и считая, что в начальный момент времени t = 0 деформация = 0. B результате получим: 
где величина  называется временем запаздывания.
называется временем запаздывания.
Скорость нарастания деформации максимальна при t = 0: 
Отсюда следует, что чем больше величина вязкой компоненты , тем меньше угол наклона кривой (t).
Деформация (t) растет с убывающей скоростью и асимптотически приближается к стационарному значению  :
: 
Таким образом, уравнение достаточно хорошо описывает процесс нарастания деформации (t), полученный в эксперименте.
В терминах механической модели (рис, 5) и модели скользящих нитей (рис. 3> параллельный упругий элемент Е1, определяет механические свойства внешних мембран клеток (сарколеммы) и внутренних структур — Т-системы и саркоплазматического ретикулума.
Последовательный элемент Е2определяет упругость актин-миозинового комплекса, обусловленную, прежде всего, местами прикрепления актина к Z-дискам и местами соединения мостиков с активными центрами тонких нитей.
 Вязкий элемент обусловлен скольжением нитей актина относительно миозина. Эта компонента резко возрастает при пассивном режиме мышцы, т. к. в этом случае мостики разомкнуты. Это проявляется в возможности сильного растяжения пассивной мышцы даже при незначительных нагрузках.
Вязкий элемент обусловлен скольжением нитей актина относительно миозина. Эта компонента резко возрастает при пассивном режиме мышцы, т. к. в этом случае мостики разомкнуты. Это проявляется в возможности сильного растяжения пассивной мышцы даже при незначительных нагрузках.
Важно, что уже в этой модели было показано существование вязкой компоненты в мышце, но  физическая природа ее оста вались не ясной.
физическая природа ее оста вались не ясной.
Активное сокращение мышцы.Для исследования характеристик сокращающихся мышц используют два искусственных режима:
1. Изометрический режим, при котором длина мышцы l = const, а регистрируется развиваемая сила F(t).
2. Изотонический режим, при котором мышца поднимает постоянный груз Р = const, а регистрируется изменение ее длины во времени  .
.
При изометрическом режиме с помощью фиксатора (рис 6. а) предварительно устанавливают длину мышцы l. После установки длины на электроды Э подается электрический стимул, и с помощью датчика регистрируется функция F(t). Вид функции F(t) в изометрическом режиме для двух различных длин представлен на рис. 7, а.
Максимальная сила Рп, которую может развивать мышца, зависит от ее начальной длины и области перекрытия актиновых и миозиновых нитей, в которой могут замыкаться мостики: при начальной длине саркомера 2,2 мкм в сокращении участвуют все мостики (см. рис. 4).
Поэтому максимальная сила генерируется тогда, когда мышца предварительно растянута на установке (рис, 6, а) так, чтобы длины ее саркомеров были близки к 2,2 мкм. На рис. 7, а это соответствует начальным длинам двух мышц  . Но, т. к. количество мостиков в мышце
. Но, т. к. количество мостиков в мышце  , больше, чем в мышце
, больше, чем в мышце  (
( 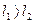 ), то сила, генерируемая 1г больше.
), то сила, генерируемая 1г больше.
При изотоническом режиме к незакрепленному концу мышцы подвешивают груз Р (рис.6,б). После этого подаётся стимул и регистрируется изменение длины мышцы во времени: . Вид этой функции в изотоническом режиме для двух различных нагрузок показан на рис. 7.б.
Как следует из рис. 7, б, чем больше груз Р , тем меньше укорочение мышцы и короче время удержания груза. При некоторой нагрузке Р = Р0 мышца совсем перестает поднимать груз; это значение. Р0 и будет максимальной силой изометрического сокращения для данной мышцы (рис. 7. а).
Здесь важно отметить, что при увеличении нагрузки угол наклона восходящей части кривой изотонического сокращения уменьшается (рис. 7,б): 
Биомеханические факторы, определяющие спортивную работоспособность
Вязко-упругие свойства мыщц разгибателей голеностопного сустава
В 1970 г. Д. Кавагна опубликовал метод определения вязко-упругих свойств икроножной мышцы человека. Испытуемые должны были выполнять небольшой прыжок вверх (20–25 см) с приземлением на вытянутые носки, при этом колени не сгибаются, руки прижаты к выпрямленному телу; испытуемый должен удерживаться на носках в течение 2–3 с.
Прыжки выполняются на динамометрической платформе. Сила, измеряемая на платформе, обусловлена действием сил тяжести и инерции:

где P = m × g — вес испытуемого, a — ускорение ОЦМ испытуемого, g — 9,81 м×с -2 .
При выполнении прыжков были получены динамограммы с колебанием показателя силы около линии веса. Эти колебания выглядели как затухающие, что и послужило основой для предположения о моделировании тела в виде пружины, массы и вязкого демпфера.
Уравнение движения демпфированного осциллятора имеет вид:

Интегрирование дает следующее решение:

где δ — фазовая константа, ω = 2πt — угловая скорость.
Когда cos (ωt + δ) равен +1 или −1 (момент достижения динамограммой максимума):
 ,
,двойное дифференцирование выражения и последующее логарифмирование дает:

или

откуда

Если выполнить измерения на одной осциллограмме: время появления первого пика после пересечения линии веса t1, F1/m−g и время появления второго пика силы t2, F2/m−g, то можно вычислить величину коэффициента демпфирования:

Коэффициент жесткости находится из выражения

отсюда получаем:

где  (частота колебаний массы).
(частота колебаний массы).
Пример: пусть t1 = 0,05 c, F1 = 2250 H, t2 = 0,400 c, m = 72 кг, F2 = 1150 H, тогда В = 447 Н/м/с, К = 2,58×10 4 Н/м.
Эта оценка жесткости в 0,66 раза меньше величины, полученной самим Кавагна (3,8×10 4 Н/м), но совпадает с оценками, полученными другими исследователями в аналогичном тесте 2,67 ± 0,48×10 4 Н/м (см. В. М. Зациорский, в соавт., 1981).
Поскольку наши расчеты были выполнены по данным Кавагна, то предполагаем, что им была допущена ошибка в расчетах.
Коэффициент жесткости, полученный таким образом, был пересчитан в коэффициент жесткости для мышц (с учетом плеч действия сил) и составил 13×10 4 Н/м. Эта оценка оказалась значительно меньше, чем величины, полученные при прямых измерениях на различных мышцах человека. При силе тяги одной мышцы 2250 Н коэффициент жесткости должен быть равен 20×10 4 или 25×10 4 Н/м, т. е. в 2,0 раза больше, чем по методике Кавагна. Зависимость «жесткость—сила» имеет линейный вид и может быть представлена следующей формулой К = 125 × F.
Заниженная величина коэффициента жесткости свидетельствует об ошибочности исходных посылок методики Кавагна. Это следует из данных Хилла (1972).
Эффект затухания колебаний активной мышцы нельзя объяснять изменением вязкости мышцы при ее переходе от состояния покоя к активности . «Все, что известно в настоящее время о мышце, говорит о бессмысленности вязко-упругой теории и модели; лучше всего забыть о них» (Хилл, 1972).
Можно предположить, что по мере растягивания активной мышцы часть мостиков или миофибрилл (при больших напряжениях) разрушается, что и приводит к потере ЗУД.
В чем причина ошибки метода Кавагна?
В рамках программно-целевого подхода ошибочность подхода Кавагна обнаруживается легко. Им был нарушен один из основополагающих принципов научного познания — принцип природной специфичности. Нельзя было по методу аналогии заменять мышцу объектом (моделью), не имеющим сходства по строению и свойствам элементам реальной мышцы.
Попробуем построить более правдоподобную модель объекта — человека, выполняющего описанный выше прыжок.
Человек может быть смоделирован системой, состоящей из трех элементов: первый — туловище и ноги, второй — стопы и третий — мышца.
Свойства элементов модели
Предполагаем, что первый и второй элементы — абсолютно твердые тела, соединенные идеальным цилиндрическим шарниром (без трения). Стопа не имеет массы, а центр масс туловища расположен на высоте Н (100 см) от оси.
Свойства и модель мышцы. Представим мышцу в виде модели, состоящей из сократительного и упругого элементов. Последовательный упругий элемент модели представляет упругие свойства сухожилий и миофибрилл мышечных волокон. Увеличение числа активированных мышечных волокон означает прибавление количества упругих элементов — рост жесткости активной мышцы.
Сократительные компоненты модели функционируют в соответствии с теорией Дещеревского (1968). Взаимодействие между актином и миозином осуществляется посредством мостиков. Мостики находятся в трех состояниях: замкнутые мостики, развивающие тянущую силу; замкнутые мостики, тормозящие скольжение нитей; разомкнутые мостики. Мостики замыкаются независимо друг от друга и тянут нити, вызывая активное сокращение.
Далее те же мостики тормозят движение, так как нити переместились и, наконец, мостики разрываются. Обозначив через n0 полное число активных центров для образования мостиков, через n — число тянущих и m — число тормозящих мостиков, получаем:

Первое слагаемое — скорость образования мостиков, второе слагаемое — скорость образования тормозящих мостиков.


Второе слагаемое — скорость размыкания мостиков. Здесь V/L — константа скорости превращения тянущих мостиков в тормозящие, равная скорости относительного перемещения нитей (V), деленной на расстояние (L) между двумя соседними активными центрами актина, на которых может быть замкнут мостик. К этим уравнениям добавляется выражение второго закона Ньютона:
где m — перемещающая масса, fo — активная сила, развиваемая одним тянущим мостиком, f — внешняя сила (нагрузка) приходящаяся на один мостик. Уравнение описывает движение с трением, выраженным как fo × m (а не вязкость мышцы).
В стационарных условиях n = m = 0 и v = 0 получаем

Это уравнение совпадает с уравнением Хилла. Таким образом, эмпирическое уравнение Хилла выведено теоретически. Уравнение отвечает стационарному скольжению нитей с силой трения, пропорциональной скорости (теория цитируется по И. В. Волькенштейну, 1981).
В случае, когда внешняя сила превышает тянущую силу мостиков, сократительный компонент остается в напряженном состоянии. Увеличение внешней силы приводит лишь к удлинению упругих компонентов мышцы, мостики остаются сцепленными, а значит нет затрат энергии (АТФ) на преодоление внешнего сопротивления. При достижении силы, вызывающей резерв сцепленных мостиков, упругие элементы миофибриллы приходят в исходное (ненапряженное) состояние, а энергия упругой деформации рассеивается, превращается в тепло. Поскольку активная мышца не обладает вязкостью, то «сокращение» упругих элементов происходит практически мгновенно (масса мышцы мала) и уже через 1 мс появляются новые мостики. Эти мостики могут преодолеть действие внешней силы, но сначала будут растянуты упругие элементы мышцы в соответствии с действующей силой.
Количество образующихся в мышце мостиков зависит от концентрации ионов в мышечных волокнах. После начала активации мышцы тянущее усилие появляется на мгновение, а только через 25–50 мс достигает максимума, а камбаловидной (медленной мышцы) до 100 мс (Коц, 1982). Развитие активного состояния связывают с увеличением концентрации ионов кальция в саркоплазме мышечных волокон.
Пока активность мышцы растет (увеличивается концентрация кальция) в сократительном компоненте внешнюю нагрузку преодолевают те мостики, которые были образованы до начала растягивания. Одновременно образуются новые мостики, но они не могут взять на себя нагрузку, поскольку им нечего преодолевать. Однако в случае разрыва уже растянутых мостиков появляется нагрузка на образовавшихся мостиках. Заметим, что при разрыве мостиков теряется энергия. Если новых мостиков не образуется, то после разрыва тех, которые преодолевали внешнее сопротивление в мышце, должно снизиться напряжение.
Модель голеностопного сустава
Модель ГСС включает элементы: стопа, туловище, две мышцы. Стопа и туловище являются абсолютно твердыми телами — безинерционными. Они соединены цилиндрическим шарниром без трения. На конце туловища расположена масса.
Мышцы — эквивалентные разгибатели стопы. Первая мышца заменяет (моделирует) трехглавую мышцу голени, вторая мышца заменяет: заднюю, большеберцовую, длинный сгибатель пальцев, длинный сгибатель большого пальца, длинную малоберцовую, короткую малоберцовую.
Первая мышца М-1 крепится на пятке (плечо тяги мышцы L1 = 4 cм в рамках подвижности сустава 60° не меняется).
Вторая мышца имеет другое плечо тяги, L2 она сгибает цилиндрический сустав, поэтому плечо не меняется и составляет 0,2 × L1 = L2 = 0,8 cм.
Предположим, что в ходе движения:
- ОЦМ находится над точкой касания стопы поверхности,
при повороте стопы происходит подъем ОЦМ по радиусу R — 18 см.
Поэтому, если L1 = 4 см, то поворот стопы, обеспечивающий подъем ОЦМ на 10 см, вызовет сокращение мышцы I:

Ускорение мышцы 2 составит
- ΔL2 = ΔL1 × 0,2 = 0,44
Cледствие
Подъем на 10 см ОЦМ соответствует предельной подвижности голеностопного сустава, дальнейший подъем до плюс 2 см будет связан с деформацией тканей и затем упором костей стопы и голени друг в друга, поэтому 2,2 см + 3 см = предельная величина сокращения М-1, которая согласуется с оценкой растяжения упругой компоненты мышцы 2 см (именно трехглавой, по Александеру, 1977).
Поворот стопы на приводит к уменьшению плеча действия силы реакции опоры, так как вращение идет не вокруг оси, а путем качения — центр давления и ось вращения смещаются вперед одновременно с центром вращения ГСС. Следовательно,
- R = const = 14 см
Сила тяги М-1 раскладывается на две составляющие: вдоль пяточной кости и перпендикулярную к ней. Последняя составляющая вызывает момент

В рамках данной модели R = 9 см, поэтому

Следовательно, при сделанных нами допущениях о конструкции модели ГСС можно утверждать, что при подъеме на носки между силой тяги мышцы М-1 и силой реакции опоры остается постепенное соотношение — сила тяги мышцы в два раза превышает силу реакции опоры.
Так как LR = const, то сила тяги мышцы М-2 может быть вычислена

Основные параметры мышц. Мышца М-1 является эквивалентной, заменяет икроножную и камбаловидную. Предположим, что имеет: физиологический поперечник — 284 см 2 , 2 ноги;
максимальную силу тяги — 11399 Н, 2 ноги;
угол перистости — 15°;
медленные мышечные волокна — 70 %;
быстрые мышечные волокна — 30 %.
Мышца М-2 является эквивалентной, заменяет заднюю большеберцовую, длинную и короткую, малоберцовые, сгибатели пальцев и большого пальца.
Предположим, что М-2 имеет: физиологический поперечник — 147 см 2 , 2 ноги;
максимальную силу тяги — 5800 Н, 2 ноги;
угол перистости — 15°;
медленные мышечные волокна — 45 %;
быстрые мышечные волокна — 55 %.
Оценка параметров выполнена по данным Pierrynowck, 1982.
Адекватность модели
После построения модели необходимо проверить способность ее прогнозировать явления, наблюдаемые в реальном объекте. С этой целью были выполнены четыре эксперимента.
Испытуемый (масса 87 кг) вставал на носки на тензоплатформе четыре раза с разной степенью активации мышц: очень медленно, со средней интенсивностью, быстро, максимально быстро. Регистрировалась реакция опоры. Запись велась на светолучевом осциллографе, скорость протеина ультрафиолетовой бумаги 250 мм/с. Тензоплатформа конструкции ВИСТИ, частота собственных колебаний более 500 ГСС (все последующие эксперименты были выполнены на этой методической базе).
Полученные динамограммы были обработаны с использованием процедуры численного интегрирования. Исходные формулы:


По данным эксперимента был рассчитан градиент силы:

где ΔR — прирост силы реакции опоры от линии веса до максимума; Δt — интервал времени, за которое произошло увеличение силы.
Анализ данных показал, что с увеличением степени активации мышц разгибателей ГСС растет пик силы реакции опоры, линейно увеличивается градиент силы.
Очевидно, в рамках нашей модели мышцы увеличение градиента силы связано с увеличением количества рекрутированных ДЕ (активных мышечных волокон). Градиент J — скорость нарастания напряжения в мышце — зависит от числа активированных МВ, следовательно, удельный градиент должен быть константой.
В формуле:

Постоянный множитель 13,3 является такой константой, характеризующей предельные возможности развития напряжения в МВ во времени.
Эта константа должна зависеть от:
- — скорости выхода кальция из саркоплазматического ретикулума;
— АТФ-ной активности миозима (мышечной композиции);
— длины мышечного волокна (быстрее растягивается ПОУК мышцы).
Следовательно,

Должен характеризовать индивидуальные особенности мышц разгибателей ГСС, связанные со скоростью развития напряжения в «среднем» МВ этих мышц. Этот показатель может служить в качестве критерия для классификации спортсменов на быстрых и медленных (по мышце , трехглавой мышце голени это замечание станет понятно из написанного ниже).
Следующий результат эксперимента. Оказалось, что падение силы реакции опоры наблюдается при одинаковой скорости подъема ОЦМ и сокращения мышцы.
Таблица 1. Путь, скорость и сила реакции опоры при вставании на носки с разной интенсивностью
| № | Интенсивность | Путь ОЦМ, м | Скорость ОЦМ, м/с | Сила, H | Скорость мышц | Мощь, Вт | |
|---|---|---|---|---|---|---|---|
| М-2 | |||||||
| 1 | Медленно | 0,049 | 0,46 | 1150 | 0,23 | 0,046 | 529 |
| 2 | Средне | 0,033 | 0,48 | 1280 | 0,24 | 0,048 | 614 |
| 3 | Быстро | 0,025 | 0,44 | 1408 | 0,22 | 0,044 | 619 |
| 4 | Максимально | 0,018 | 0,44 | 1568 | 0,22 | 0,044 | 689 |
Эта скорость для ОЦМ составила 0,44 м/с, для — 0,22 м/с, для М-2 — 0,044 м/с.
Угловая скорость разгибания стопы в момент пика силы достигла 250 град/с при выполнении упражнения с максимальной интенсивностью, в этот момент мощность разгибания достигла 689 Вт.
При вставании на носки испытуемый поднялся на 10 см (максимальная активность мышц), следовательно, выполнил работу:

Испытуемый массой в 87 кг выпрыгнул вверх и приземлился на носки напряженных ног.
Испытуемый выпрыгнул на 0,159 м. К моменту касания опоры имел скорость ОЦМ 1,77 м/с.
Энергия упругой деформации накапливалась в течение 100 мс. За это время ОЦМ опускался на 11 см, что соответствует удлинению на 2,2 см. На динамограмме отмечено наложенное колебание силы с частотой 40–50 Гц. При удлинении с 2,0 см до 2,2 см произошло падение силы реакции опоры, рассеивание энергии. Подъем ОЦМ составил 5,6 см.
За время амортизации испытуемый выполнил следующую работу:

За время возвращения в исходное состояние («отталкивания»):

Таким образом, за время амортизации было рассеяно 186 Дж.
Если предположить, что подъем ОЦМ был выполнен за счет ЗУДМ, то можно рассчитать коэффициент жесткости. Cледовательно, работа могла быть выполнена за счет ПОУК мышц разгибателей голени:

Сопоставление реально выполненной работы (47 Дж) с энергией, накопленной в мышцах (82 Дж), показывает, что при разгибании ГСС наблюдается потеря энергии — КПД 57%.
Испытуемый (m = 87 кг) выпрыгивал и приземлялся на носок одной выпрямленной ноги.
Скорость в момент начала амортизации составила 1,23 м/с, высота падения ОЦМ 9,6 см. Амортизация длилась 140 мс. За это время ОЦМ опустился на 13,7 см. Следовательно, была выполнена работа A = 87 × 9,81 × 0,233 = 199 Дж.
За время отталкивания была выполнена работа:
- A = mqh = 87 × 9,81 × 0,07 = 60 Дж
Коэффициент жесткости составил:

Работа за счет энергии, накопленной в ПОУК, может составить:

Сопоставление выполненной работы с ЗУДМ показало, что часть энергии рассеивается — КПД 62,5 %.
Прыжки вверх на носках без сгибания ног. Испытуемый m=87 кг. Скорость ОЦМ при приземлении составила 1,6 м/с, при вылете 1,56 м/с. За время взаимодействия с опорой была выполнена работа в фазе амортизации или в фазе отталкивания:

Время амортизации опускания ОЦМ — 80 мс, отталкивания, подъема ОЦМ — 80 мс. Максимум силы реакции опоры составил 5000 Н. Так же, как в эксперименте 2, были зарегистрированы наложенные колебания с периодом 20—25 мс в ходе амортизации.
Рассчитаем коэффициент жесткости:

Следовательно, энергия, запасенная в ПОУК, составит:

В отличие от двух предыдущих упражнений ЗУЦМ не хватило для предсказания выполненной механической работы (различие 20 Дж). Недостающие 20 Дж могут добавить мышцы, входящие в группу М-2.
Обсуждение результатов исследования
Цель этой статьи — изучить проявление вязких и упругих свойств мышц разгибателей голени. Ответ на вопрос о наличии вязких свойств у мышц был дан Хиллом, ответ, как известно, был отрицателен: нет вязкости, во всяком случае той, которая получается по оценке методики Кавагны или Хилла в годах. Что же касается упругости, то в трех экспериментах 2, 3, 4, где мышцы функционировали в уступающем режиме, а затем в преодолевающем, были получены неоднозначные результаты.
- Сопоставим величины коэффициентов жесткости. Во втором эксперименте

в третьем —

в четвертом —

Из этих чисел видно, что коэффициенты жесткости оказались сходными, для двух ног:

для одной:

Для случая, когда активная только эквивалентная мышцы коэффициент жесткости составит:
- для двух ног:
 ;
; для одной ноги:
 .
.Коэффициенты жесткости, полученные в наших экспериментах, хорошо согласуются с данными литературы (см. В. М. Зациорский с сотр., 1981). На этом можно было бы остановиться, однако обнаруженное нами явление рассеивания энергии в тестах с удержанием позы и отсутствием рассеивания при выполнении прыжков на носках требует объяснения.
Для объяснения этого явления воспользуемся описанной выше моделью голеностопного сустава. Адекватность этой модели был подтверждена результатами первого эксперимента. Например, независимо от степени активации (количества рекрутированных двигательных единиц) максимальная сила и последующее снижение величины опорной реакции наблюдается при одной и той же скорости. Это вызвано тем, что все рекрутированные ДЕ функционируют с максимальной интенсивностью, активация МВ идет одинаково, а увеличение внутреннего трения (числа тормозящих мостиков) зависит только от скорости, видимо, существует индивидуальный оптимум, при котором наблюдается максимум проявляемой силы. Для этот оптимум соответствует 0,22 м/с, для М-2 этот оптимум должен быть выше, так как мышцы, входящие в этот «эквивалент», более быстрые.
Поэтому при выполнении максимальных прыжков вверх или в беге сократительный компонент М-2 должен выполнять работу, а в положительную работу может совершить только упругий компонент.
При выполнении прыжков на носках испытуемый в каждом прыжке выполнял отрицательную и положительную работу. В том прыжке, который был зарегистрирован, она составила по 181 Дж. Исходя из данных изменения высоты подъема ОЦМ на опоре (0,08 м) и коэффициента жесткости мышц разгибателей (50×10 4 Н/м) была вычислена работа, связанная с накоплением и отдачей энергии упругой деформации мышц. Она составила 160 Дж.
Это означает, что основная доля работы приходится на ПОУК. В фазе амортизации была положена энергия в ПОУК 160 Дж, а 21 Дж рассеялся (КПД = 88,4 %). В фазе отталкивания 160 Дж было возвращено ПОУК, а 21 Дж был компенсирован М-2, ее сократительным компонентом. Напомним, что в момент ухода с опоры скорость ОЦМ составила 1,48 м/с, на этой скорости сократительный компонент работы выполнить не может (468 градусов/с), а сократительный компонент М-2 будет иметь скорость укорочения 0,148 м/с, что значительно меньше найденной в первом эксперименте, «оптимальной» скорости — 0,22 м/с.
Если предположить, что в четвертом эксперименте наши рассуждения правильные, то результаты второго и третьего экспериментов доказывают некорректность наших рассуждений и соответственно неадекватность модели.
Расхождение в экспериментальных данных можно объяснить лишь введением в модель дополнительного элемента — мышцы антагониста (м-3). Если мы введем предположение, что для выполнения задания по удержанию заданной позы (приземлиться и остаться на вытянутых носках стоп) необходимо одновременно активировать как мышцы агонисты (, М-2), так и антагонисты (М-3), то результаты и экспериментов становятся понятны.
Физиологический поперечник мышц группы М-3 составляет:
- передняя большеберцовая — 39,5 см 2
длинный разгибатель пальцев — 26,5 см 2
длинный разгибатель большого пальца — 12,9 см 2
Итого: 78,9 см 2
Следовательно, М-3 может проявить изометрическую силу 78,9 × 40 = 3156 Н. Для двух ног 6300 Н.
Эта сила может создавать момент в голеностопном суставе (плечо тяги 0,02 см) 3156 × 0,06 = 189 Н × м.
- Момент = 11399 × 0,04 = 456 Н × м
Момент М-2 = 5800 × 0,015 = 87 Н × м
Следoвательно, если будут одновременно активные все три эквивалентные мышцы, то в уступающем режиме в фазе амортизации доля силы М-3 будет неощутима. Поэтому наши оценки коэффициентов жесткости мышц, вычисленные по силам, возникшим при максимальном растяжении ПОУК, оказались одинаковыми. Иная ситуация возникает при подъеме ОЦМ (фаза отталкивания). В этом случае М-3 работает в уступающем режиме. Выполним работу, необходимую для растяжения М-3:

Здесь К — коэффициент жесткости, Х — удлинение М-3 при повороте на 0,5 радиана.
Коэффициент жесткости М-3 был оценен следующим образом: сила изометрической тяги должна быть равна 22399, при этом Кж = 50 × 10 4 , тогда для М-3 = 6300 Н.

Следовательно, при растягивании активной М-3 должна быть выполнена работа 60—100 Дж (если удлинение составляет 3—4 см).
Таким образом, получаем следующий баланс в выполненной работе:
- Эксперимент 2. За время амортизации было накоплено около 82 Дж энергии, в ПОУК порядка 150 Дж было рассеяно в результате разрыва мостиков и миофиламентов. При выполнении отталкивания ЭУД ПОУК пошло на преодоление сопротивления М-3 (30–40 Дж), на устранение ОЦМ (10–12 Дж) и подъем ОЦМ (30–40 Дж).
Эксперимент 3. За время амортизации было накоплено 96 Дж. Около 100 Дж было рассеяно. За время отталкивания 60 Дж пошло на преодоление внешних сил (инерции и тяжести) и 30–40 Дж на растягивание ПОУК М-3.
Из представленных расчетов можно видеть, что поведение модели адекватно реальному объекту в случае приближения ее устройства к реальной действительности (с более точным учетом природной специфичности).
Далее для достижения цели, удержания заданной позы целесообразно активировать мышцы агонисты, так и антагонисты. В этом случае система становится более жесткой, амплитуда колебаний быстрее уменьшается.
Результаты, полученные в нашем исследовании, противоречат выводам, сделанным в работе Schenau c cоавт. В их работе было показано, что сократительный компонент мышц разгибателей ГСС не может выполнить работу 191 Дж и создать максимальную мощность 4000 Вт. По оценкам этих авторов, не может выполнить такой работы и ПОУЭ этих мышц, поскольку максимум, что он может дать 50–100 Дж. В связи с этим было введено предположение, что при выполнении волейбольного прыжка голеностопный сустав выполняет работу 190 Дж и достигает мощности 4000 Вт, благодаря передаче мощности от коленного сустава к ГСС через двухсуставную икроножную мышцу.
Видимо, не следует отрицать возможность передачи энергии по двухсуставным мышцам, однако нельзя согласиться с заниженными оценками накопления ЭУД и ПОУК мышц разгибателей голени.
Результаты нашего исследования убедительно показали, что мышцы разгибатели ГСС могут накапливать в ПОУК 160–200 Дж и выполнять преодолевающее движение с максимальной мощностью 4000–5000 Вт.
http://megaobuchalka.ru/11/47254.html
http://mipt.ru/education/chair/sport/science/biomechanics/work-2