Пассивный перенос веществ через мембрану
Пассивный транспорт— это перенос вещества из мест с большим значением электрохимического потенциала к местам с его меньшим значением.
Пассивный транспорт идет с уменьшением энергии Гиббса, и поэтому этот процесс может идти самопроизвольно без затраты энергии.
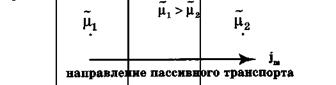
Рис.Схема пассивного транспорта
Плотность потока вещества jm при пассивном транспорте подчиняется уравнению Теорелла:
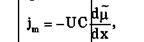
где U — подвижность частиц, С — концентрация. Знак минус показывает, что перенос происходит в сторону убывания μ.
Для разбавленных растворов при μ = const плотность потока вещества выражается уравнением Нернста-Планка:
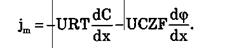
где U — подвижность частиц.
Итак, могут быть две причины переноса вещества при пассивном транспорте: градиент концентрации dC / dxи градиент электрического потенциала dφ / dx.Знаки минусов перед градиентами показывают, что градиент концентрации вызывает перенос вещества от мест с большей концентрацией к местам с его меньшей концентрацией; а градиент электрического потенциала вызывает перенос положительных зарядов от мест с большим к местам с меньшим потенциалом.
В случае неэлектролитов (Z = 0) или отсутствия электрического поля (dφ/dx =0) получаем уравнение:
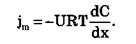
Согласно соотношению Эйнштейна коэффициент диффузии D=URT. В результате получаем уравнение, описывающее простую диффузию — закон Фика:
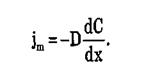 |
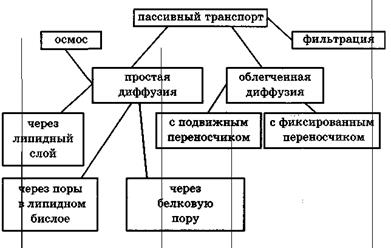 |
Рис.Классификация видов пассивного транспорта
Диффузия — самопроизвольное перемещение вещества из мест с большей концентрацией в места с меньшей концентра вещества вследствие хаотического теплового движения.
Диффузия вещества через липидный бислой вызывается градиентом концентрации в мембране.
Коэффициент проницаемости мембраны зависит от свойств мембраны и переносимых веществ.
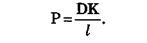
Величина К носит название коэффициента распределения, который показывает соотношение концентрации вещества вне мембраны и внутри ее. Коэффициент проницаемости тем больше, чем больше коэффициент диффузии (чем меньше вязкость мембраны), чем тоньше мембрана (чем меньше l) и чем лучше вещество растворяется в мембране (чем больше К).
Хорошо растворимы в фосфолипидной фазе мембраны неполярные вещества, например органические жирные кислоты, эфиры. Этим вещества хорошо проникают через липидную фазу мембраны.
Плохо проходят через липидный бислой полярные, водорастворимые вещества: соли, основания, сахара, аминокислоты, спирты.
В биологических мембранах был обнаружен еще один вид диффузии — облегченная диффузия. Облегченная диффузия происходит при участии молекул переносчиков. Например, валиномицин — переносчик ионов калия. Молекула валиномицина имеет форму манжетки, устланной внутри полярными группами, а снаружи — неполярными.
Молекулы валиномицина, оказавшиеся у поверхности мембраны, могут захватывать из окружающего раствора ионы калия. Диффундируя в мембране, молекулы переносят калий через мембрану, и некоторые из них отдают ионы в раствор по другую сторону мембраны. Таким образом, происходит перенос иона калия через мембрану валиномицином.
Облегченная диффузия, таким образом, происходит от мест с большей концентрацией переносимого вещества к местам с меньшей концентрацией. По-видимому, облегченной диффузией объясняется также перенос через биологические мембраны аминокислот, сахаров и других биологически важных веществ.
Отличия облегченной диффузии от простой:
· перенос вещества с участием переносчика происходит значительно быстрее;
· облегченная диффузия обладает свойством насыщения: при увеличении концентрации с одной стороны мембраны плотность потока вещества возрастает лишь до некоторого предела, когда все молекулы переносчика уже заняты;
· при облегченной диффузии наблюдается конкуренция переносимых веществ в тех случаях, когда переносчиком переносятся разные вещества; при этом одни вещества переносятся лучше, чем другие, и добавление одних веществ затрудняет транспорт других; так, из сахаров глюкоза переносится лучше, чем фруктоза, фруктоза лучше, чем ксилоза, а ксилоза лучше, чем арабиноза, и т.д.;
· есть вещества, блокирующие облегченную диффузию – они образуют прочный комплекс с молекулами переносчика, например, флоридзин подавляет транспорт сахаров через биомембрану.
Фильтрацией называется движение раствора через поры в мембране под действием градиента давления P. Скорость переноса при фильтрации подчиняется закону Пуазейля:
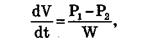
гда dV/dt — объемная скорость переноса раствора, w — гидравлическое сопротивление/
Явление фильтрации играет важную роль в процессах переноса воды через стенки кровеносных сосудов.
Осмос— преимущественное движение молекул воды через полупроницаемые мембраны (непроницаемые для растворенного вещества и проницаемые для воды) из мест с меньшей концентрацией растворенного вещества в места с большей концентрацией. Осмос — по сути дела, простая диффузия воды из мест с ее большей концентрацией в места с меньшей концентрацией воды. Осмос играет большую роль во многих биологических явлениях. Явление осмоса обусловливает гемолиз эритроцитов в гипотонических растворах.
Дата добавления: 2016-02-02 ; просмотров: 2786 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Транспорт веществ через мембраны
Пассивный транспорт.
Если вещество движется через мембрану из области с высокой концентрацией в сторону низкой концентрации (т. е. по градиенту концентрации этого вещества) без затраты клеткой энергии, то такой транспорт называется пассивным, или диффузией. Различают два типа диффузии: простую и облегченную.
Простая диффузия.
Характерна для небольших нейтральных молекул (H2O, CO2, O2), а также гидрофобных низкомолекулярных органических веществ. Эти молекулы могут проходить без какого-либо взаимодействия с мембранными белками через поры или каналы мембраны до тех пор, пока будет сохраняться градиент концентрации.
Облегченная диффузия.
Характерна для гидрофильных молекул, которые переносятся через мембрану также по градиенту концентрации, но с помощью специальных мембранных белков — переносчиков. Для облегченной диффузии, в отличие от простой, характерна высокая избирательность, так как белок переносчик имеет центр связывания комплементарный транспортируемому веществу, и перенос сопровождается конформационными изменениями белка. Один из возможных механизмов облегченной диффузии может быть следующим: транспортный белок (транслоказа) связывает вещество, затем сближается с противоположной стороной мембраны, освобождает это вещество, принимает исходную конформацию и вновь готов выполнять транспортную функцию. Мало известно о том, как осуществляется передвижение самого белка. Другой возможный механизм переноса предполагает участие нескольких белков-переносчиков. В этом случае первоначально связанное соединение само переходит от одного белка к другому, последовательно связываясь то с одним, то с другим белком, пока не окажется на противоположной стороне мембраны.
Активный транспорт.
Имеет место в том случае, когда перенос осуществляется против градиента концентрации. Такой перенос требует затраты энергии клеткой. Активный транспорт служит для накопления веществ внутри клетки. Источником энергии часто является АТФ. Для активного транспорта кроме источника энергии необходимо участие мембранных белков. Одна из активных транспортных систем в клетке животных отвечает за перенос ионов Na+ и K+ через клеточную мембрану. Эта система называется Na+ — K+ — насос. Она отвечает за поддержание состава внутриклеточной среды, в которой концентрация К+ выше, чем Na+.
Математическое моделирование процессов переноса веществ через мембрану клетки.
Свободная диффузия
Вещества, перемещающиеся через мембрану путём свободной диффузии, не образуют каких-либо химических связей с другими веществами.
Для количественной характеристики диффузии используют физическую величину — поток вещества (J) :
J=dn/dt*1/S (1),
где n — количество вещества в молях, перемещающихся посредством диффузии через поверхность S, перпендикулярную потоку вещества, за единицу времени.
Первый закон Фика указывает, что поток вещества, перемещаемого путём диффузии, пропорционален движущей силе диффузии — градиенту концентрации вещества: J = — D · dC/dx (2).
Отрицательный знак означает, что поток направлен из области высокой концентрации вещества в область с более его низкой концентрацией, в результате чего градиент концентрации уменьшается. D — коэффициент диффузии, который зависит от природы вещества и температуры:
D=U·R·T (3),
где U — подвижность частиц вещества, R — универсальная газовая постоянная, T — абсолютная температура.
Если диффузия осуществляется через мембрану, уравнение (2) может быть представлено как
J=-P·(C1 -C2) (4),
где C1 и C2 — концентрация раствора внутри и вне клетки, P — коэффициент проницаемости мембраны для данного вещества. Коэффициент проницаемости определяется коэффициентом диффузии D вещества, толщиной мембраны d и коэффициентом распределения вещества K, зависящим от растворимости вещества в органических растворителях, но не воде.
P=Dk/d (5)
Проницаемость мембраны для неэлектролитов существенно зависит от их способности растворяться в билипидном слое мембраны. Проницаемость мембраны для различных веществ определяют по растворимости в оливковом масле, которую можно рассматривать как модель мембранных липидов. Таким образом, мембрана хорошо проницаема для липидорастворимых веществ (спирты, эфиры), не имеющих биологического значения. Но такие гидрофильные вещества как сахара, аминокислоты не способны проникать через биологическую мембрану посредством свободной диффузии. Для этого требуются специальные системы транспорта (смотри ниже).
Проницаемость мембраны зависит также от размера молекул. Мелкие молекулы могут проникать через мембрану путём свободной диффузии. Например, вода не растворима в липидах и органических растворителях. Но она проникает через плазматическую мембрану благодаря небольшому размеру молекул. Проницаемость мембраны для воды очень высокая. Предполагают, что она проникает в мембрану через временные структурные дефекты, формирующихся при тепловых колебаниях хвостиков из жирных кислот. Эти дефекты (кинки) позволяют перемещаться через мембрану не только молекулам воды, но также другим небольшим гидрофильным молекулам (кислород, углекислый газ).
Облегченная диффузия
Крупные гидрофильные молекулы (сахара, аминокислоты) перемещаются через мембраны с помощью специальных молекул — мембранных переносчиков. Мембранные переносчики представляют собой интегральные белки, которые имеют центры связывания транспортируемых молекул. Образующаяся связь белка и переносчика является обратимой и обладает высокой степенью специфичности. Транспортируемая молекула проходит через мембрану вследствие изменения конформации белка-переносчика при химическом взаимодействии центров связывания обеих молекул.
Транспорт веществ через мембрану, в котором используются транспортные молекулы, называются облегчённой диффузией. Этот тип транспорта мембраны является одним из видов диффузии, поскольку транспортируемое вещество перемещается по градиенту концентрации. Никакая дополнительная энергия не требуется для этого процесса. Но облегченная диффузия отличается от свободной диффузии своей высокой специфичностью. Переносчики мембраны могут узнавать даже оптические изомеры одного и того же вещества.
Другой особенностью облегченной диффузии является феномен насыщения. Поток вещества, транспортируемого путём облегченной диффузии, растёт в зависимости от концентрации вещества только до определенной величины. Затем возрастание потока прекращается, поскольку транспортная система полностью занята. Таким образом, действие транспортной системы подобное катализу ферментами, однако переносчик не ускоряет химическую реакцию, а перемещает вещество через мембрану.
Существуют некоторые системы переносчиков, которые способны транспортировать более одного вещества. Процесс называется симпортом (или котранспортом), если вещества перемещаются в одном и том же направлении, и антипортом (встречным транспортом), если направления перемещения веществ противоположны.
Примером облегченной диффузии является действие системы транспорта глюкозы через мембраны эритроцитов и мышечных клеток. Другой пример — антипорт бикарбоната и ионов гидроксила в плазматической мембране эритроцитов.
Электродиффузия
Электродиффузия — диффузия электрически заряженных частиц (ионов) под влиянием концентрационных и электрических градиентов. Ионы — атомы или группы атомов, которые приобретают электрический заряд, теряя или приобретая электроны. Липидный бислой мембраны непроницаем для ионов. Они могут проникнуть через плазматическую мембрану только посредством специальных структур — ионных каналов, которые образованы интегральными белками.
Движущей силой диффузии является не только разность концентрации ионов внутри и вне клетки, но также разность электрических потенциалов, создаваемых этими ионами по обе стороны мембраны. Следовательно, диффузионный поток ионов определяется градиентом электрохимического потенциала (электрохимический градиент).
Электрохимический потенциал является энергией ионов:
 |
μ0— стандартный химический потенциал, который зависит от химической природы вещества и температуры, R — универсальная газовая постоянная, T — температура, C — концентрация иона, z — электрический заряд, F — константа Фарадея, φ — электрический потенциал.
Зависимость потока ионов J от электрохимического градиента определяется уравнением Теорелла:
 |
где U — подвижность ионов, C — концентрация ионов, dμ/dx — электрохимический градиент.
Подставляя (6) в (7), можно получить уравнение Нернста-Планка с учётом двух градиентов, которые обуславливают диффузию ионов:
 |
Ионные каналы мембраны представляют собой интегральные белки мембраны, которые образуют отверстия в мембране, заполненные водой. В плазматической мембране обнаружен ряд ионных каналов, которые характеризуются высокой специфичностью, допускающей перемещение только одного вида ионов. Существуют натриевые, калиевые, кальциевые и хлорные каналы. Каждый из них имеет так называемый селективный фильтр, который способен пропускать только определённые ионы. Существует несколько теорий, объясняющих избирательность ионных каналов плазматической мембраны.
Проницаемость ионных каналов может изменяться благодаря наличию ворот, определенных групп атомов в составе белков, формирующих канал. Конформационные изменения ворот переводят канал из открытого состояния в закрытое и наоборот. Механизмы регуляции положения ворот могут отличаться в различных каналах. Некоторые из них открываются при изменениях электрического потенциала мембраны. Другие открываются под действием специфических химических веществ, выполняющих сигнальные функции.
16.Активный транспорт вещества через мембрану клетки.
Активный транспорт — перенос вещества через клеточную или внутриклеточную мембрану (трансмембранный активный транспорт) или через слой клеток (трансцеллюлярный активный транспорт), протекающий из области низкой концентрации в область высокой, т. е. с затратой свободной энергии организма. В большинстве случаев, но не всегда, источником энергии для осуществления активного транспорта служит энергия макроэргических связей АТФ.
Характеристика транспортных процессов в клеточной мембране
» data-shape=»round» data-use-links data-color-scheme=»normal» data-direction=»horizontal» data-services=»messenger,vkontakte,facebook,odnoklassniki,telegram,twitter,viber,whatsapp,moimir,lj,blogger»>
ХАРАКТЕРИСТИКА ТРАНСПОРТНЫХ ПРОЦЕССОВ В МЕМБРАНЕ КЛЕТКИ
Одна из наиболее существенных функций биологических мембран – обеспечение избирательной проницаемости для веществ, транспортируемых в процессе жизнедеятельности как из клетки в среду, так и из среды во внутреннее пространство клетки. Отдельные части живой системы, разделенные мембраной, будь то разные клетки или отдельные компартменты внутриклеточной среды, функционируют как открытые системы. С помощью транспортных систем осуществляется регуляция объема клеток, величины рН и ионного состава цитоплазмы.
Среди многообразных явлений, протекающих в клетке, важное место занимают активный и пассивный транспорт веществ, осмос, фильтрация и биоэлектрогенез. Благодаря транспортным системам клетки накапливают метаболиты, важные для обеспечения энергетического цикла и метаболических процессов, выводят в окружающую среду токсические вещества, а также создают разность потенциалов на мембране. В настоящее время, очевидно, что все эти явления, так или иначе определяются барьерными свойствами клеточных мембран.
В зависимости от потребностей клетки транспорт веществ осуществляется или по или против концентрационного градиента. Способ проникновения через мембрану в значительной степени определяется свойствами вещества. Низкомолекулярные нейтральные вещества, такие, как газы, вода, аммиак, глицерин и мочевина, свободно диффундируют через биомембраны. Однако с увеличением размера молекулы теряют эту способность. К примеру, клеточные мембраны непроницаемы для глюкозы и других сахаров. Проницаемость биомембран зависит также от полярности веществ. Неполярные вещества, такие, как бензол, этанол, диэтиловый эфир и многие наркотики, способны проникать в клетку в результате диффузии. Напротив, для гидрофильных, особенно заряженных веществ, мембрана непроницаема. Однако во многих случаях именно такие вещества необходимы для функционирования клетки, поэтому в живых системах эволюционно сформировались специализированные транспортные системы для переноса таких веществ через мембрану.
Выделяют 5 разновидностей мембранного транспорта:
- пассивная диффузия;
- облегченная диффузия;
- первично-активный транспорт;
- вторично-активный транспорт;
- механизм, сопряженный с изменением структурной целостности мембран (рис. 40).

Перенос веществ через биологические мембраны осуществляется с помощью различных механизмов и протекает в основном без нарушения структурной целостности мембран. Если транспорт сопровождается уменьшением свободной энергии, он протекает самопроизвольно и называется пассивным.
Пассивный транспорт (пассивная или облегченная диффузия) происходит по направлению градиентов химического электрохимического) потенциала, результатом чего является уменьшение градиентов концентраций, если нет других процессов, которые обеспечивают их поддержание на постоянном уровне. Кроме указанных видов транспорта в мембране имеют место также процессы, не связанные со специфическими структурами. К таковым относится еще один вид транспорта.
Неспецифическая диффузия является процессом, не связанным со специфическими структурами – она осуществляется благодаря физико-химическим свойствам липидного бислоя. Объяснение механизма неспецифической диффузии через биологические мембраны малых молекул, таких как диметилформамид, вода или гидрофобные молекулы метанола, основано на представлении о динамических свойствах бислоя. Полагают, что малые молекулы могут достаточно быстро проникать через бислой вместе с кинками, возникающими в области дефектных зон в структуре бислоя. Такие короткоживущие мигрирующие образования нельзя считать порами, поскольку они не требуют специальных белковых образований. Неспецифическая диффузия осуществляется без участия специальных механизмов: вещества проникают через мембрану благодаря наличию кинков (от англ. kink – петля) или в области мембранных дефектов. Вследствие теплового движения хвостов молекул фосфолипидов кинки могут перемещаться поперек мембраны и переносить попавшие в них мелкие молекулы, в первую очередь, молекулы воды.
Облегченная диффузия требует для своего протекания наличия белков – порообразователей или переносчиков. Такими переносчиками служат антибиотики-каналоформеры (они различаются по специфичности: валиномицин высокоспецифичен для калия, иономицин – для кальция, грамицидин относительно мало специфичен для натрия) или белки порины, образующие поры для воды в наружной клеточной мембране (аквапорины) или в мембранах митохондрий (через них могут проникать даже низкомолекулярные белки).
Активный транспорт – процесс переноса веществ или ионов против их концентрационных градиентов, который так или иначе обеспечивается энергией метаболических процессов. Активный транспорт бывает первично-активным и вторично-активным. В случае ионного транспорта, обеспечиваемого транспортными АТФазами (их называют также ионными насосами), энергодающей стадией является гидролиз АТФ. Такой процесс называется первично-активным транспортом. Если же энергия обусловлена градиентом ионов, созданным независимо, например, в процессе первично-активного транспорта, этот процесс называется вторично-активным транспортом. Специальные мембранные структуры обеспечивают пассивный и активный транспорт, в число которых входят каналы, переносчики и ферменты, осуществляющие перемещение ионов (веществ) против их концентрационного градиента.
Кроме этих основных видов транспорта имеются специальные виды переноса веществ через мембрану, выделяемые в особую группу: они сопряжены с изменением структурной целостности мембран. Сюда относятся процессы высвобождения медиаторов при возбуждении синапсов (происходит слияние синаптических пузырьков с мембраной и высвобождение их содержимого в синаптическую щель), перенос генетического материала через ядерную мембрану, процессы пиноцитоза и экзоцитоза, а также деятельность пермеаз бактерий, обеспечивающих перемещение через клеточную стенку олигопептидов.
Перенос белков от одних органелл к другим происходит с помощью везикул. Везикулы отпочковываются от мембран одной органеллы, а затем исчезают, сливаясь с мембраной другой органеллы. Белки при этом находятся в полости пузырька или в составе мембран подобно интегральным белкам. Перенос через биомембраны белков со значительной молекулярной массой осуществляется специальными транспортными системами. Из цитоплазмы в ядро белки попадают через крупный (125000 кДа) заполненный водой пориновый комплекс. Транспорт белков через него энергозависим и поэтому может регулироваться. Ядерные белки несут одну или несколько сигнальных последовательностей, с помощью которых они связываются с пориновым комплексом и импортируются с сохранением третичной структуры. Импорт белков из цитоплазмы в органеллы осуществляется белками-переносчиками, которые представляют собой белковые комплексы, переносящие линейные полипептиды через биомембраны также энергозависимым образом. Специфичность процесса обеспечивается за счет связывания сигнальной последовательности с ближайшим рецептором. Процессы развертывания и вторичной укладки белков контролируются белками шаперонами.
Пассивный и активный виды транспорта ионов обеспечиваются специальными структурами. В их числе каналы, переносчики и ферменты, осуществляющие перемещение специфических ионов против их концентрационного градиента за счет энергии АТФ. Зачастую одни и те же механизмы могут использоваться и для пассивного, и для активного переноса, только в последнем случае они соединены с «биологическим трансформатором» энергии (АТФазой), обеспечивающим процесс активного транспорта энергией (рис. 40). Как пассивный, так и активный транспорт, в общем виде, подчиняется кинетике насыщения, что свидетельствует о наличии конечного числа участков переноса.
Движущими силами пассивного транспорта веществ через биологическую мембрану могут служить следующие градиенты: – концентрационный – для нейтральных молекул, – электрохимический – для ионов, – градиент гидростатического давления и осмотический градиент – для воды.
Трансмембранный градиент концентраций молекул различных веществ периодически возникает (или постоянно существует) на клеточной мембране в процессе жизнедеятельности. При определенных условиях через мембрану клетки осуществляется перемещение этих соединений в направлении их концентрационного градиента.
Следующие отличия характерны для облегченной диффузии:
- перенос ионов с участием переносчика происходит значительно быстрее по сравнению со свободной диффузией;
- облегченная диффузия обладает свойством насыщения – при увеличении концентрации с одной стороны мембраны плотность потока вещества возрастает лишь до некоторого предела, когда все молекулы переносчика уже заняты (рис. 41);
- при облегченной диффузии наблюдается конкуренция переносимых веществ в тех случаях, когда одним переносчиком переносятся разные вещества; при этом одни вещества переносятся лучше, чем другие, и добавление одних веществ затрудняет транспорт других;
- вещества, которые образуют прочный комплекс с молекулами переносчика, препятствуя дальнейшему переносу, блокируют облегченную диффузию
Каналы, сформированные из матричных белков, значительно облегчают процесс переноса, но в отличие от белков-переносчиков, пронизывая мембрану насквозь, являются неподвижными.
Канальные белки образуют в мембране заполненные водой поры, проницаемые для определенных ионов. Например, имеются специфические ионные каналы для ионов Na + , K + , Са 2+ . В отличие от ионных каналов белки-переносчики образуют специфический комплекс с молекулой субстрата, который обладает способностью легко проникать через мембрану. В этом отношении белки-переносчики (транспортные белки, пермеазы) похожи на ферменты. Единственное различие состоит в том, что они «катализируют» направленный транспорт, а не ферментативную реакцию. Они проявляют высокую специфичность – иногда групповую – к субстратам, подлежащим переносу через мембрану. Они характеризуются определенным сродством, выражаемым в виде константы K а скоростью транспорта V.

Рис. 41. Зависимость скорости транспорта от концентрации транспортируемых молекул при канальном (I) и переносчиковом (II) механизме А – схема, Б – кинетическая зависимость.
Канальный и переносчиковый типы транспорта легко различить по концентрационной зависимости скорости транспортного процесса: канальный тип имеет линейную зависимость, а переносчиковый тип характеризуется кинетикой насыщения (рис. 42).

Ионные каналы играют очень важную роль в функционировании клеток. В частности, они обеспечивают передачу нервного импульса и мышечное сокращение. При исследовании бактерий Streptomyces lividans в 1998 г. Р. Маккиннон (лауреат Нобелевской премии по химии за 2003 год) показал, что калиевый канал состоит из белков, образующих тело канала, и «ионного фильтра», который в каждый момент времени занят двумя ионами калия. Размеры фильтра точно соответствуют размерам ионов калия, вследствие этого происходит их достаточно прочное связывание. Ионы проходят через канал по эстафетному механизму, когда связывание третьего иона в фильтре приводит к высвобождению первого иона с противоположной стороны канала.
Описанный механизм позволяет совместить высокую скорость с высокой избирательностью работы канала. Выявлены структурные особенности регуляции ионных каналов, которые обеспечивают их открывание и закрывание. За это свойство отвечает расположенная на противоположной стороне от ионного фильтра часть молекулы белка, которая имеет возможность осуществлять механические движения в ответ на приложение трансмембранной разности электрических потенциалов или связывание лиганда. Долгое время считалось, что для диффузии воды через клеточные мембраны достаточно ее естественной проницаемости через липидную часть мембран за счет движения кинков. В 1988 г. в лаборатории П. Агре (лауреата Нобелевской премии по химии за 2003 год) были описаны аквапорины – новый класс белков, которые высокоэффективно пропускают молекулы воды, будучи абсолютно непроницаемы ни для каких ионов, включая протоны.
В отличие от ионных каналов, аквапорины осуществляют избирательное пропускание воды через мембраны клеток. Аквапорины имеют молекулярную массу
30 кДа и находятся в мембране в виде тетрамеров (рис. 43). Они встречаются в клетках всех живых организмов и играют особенно важную роль в физиологии почек (у человека через них проходит до 170 л воды в сутки). Нарушения работы аквапоринов (например, в случае генетических дефектов этих белков) приводят к тяжелым патологиям.

Рентгеноструктурный анализ аквапорина показал, что его структура сильно отличается от структуры калиевого канала. В мембране формируется очень узкое отверстие, в центре которого имеются два положительных заряда, расположенных на двух симметричных петлях с характерной последовательностью -N-P-A. Прохождение большинства катионов и анионов через данный канал невозможно из-за его малого размера, а протоны не проходят через него из-за наличия положительного заряда.
Примером канального механизма переноса являются натриевые каналы возбудимых мембран. Иллюстрацией транспорта переносчикового типа является АТФ/АДФ-транслоказа, обеспечивающая перенос нуклеотидов через митохондриальую мембрану. Другой пример мембранных транспортеров представляют ионофоры – гидрофобные вещества, образующие с катионами жирорастворимые комплексы, способные встраиваться в мембрану. Эти молекулы могут функционировать по принципу как подвижных переносчиков, так и каналоформеров. Принцип действия ионофоров заключается в том, что они экранируют заряд транспортируемого иона, что позволяет ему пересекать по концентрационному градиенту гидрофобную область липидного бислоя биологической мембраны.
Примером ионофора, работающего как подвижный переносчик, может служить уже упоминавшийся депсипептид (пептид, который наряду с амидными участками содержит также и сложноэфирные связи) – валиномицин, продуцируемый одним из штаммов Streptomyces. Он способен образовывать комплексы с калием на 3 порядка более активно, чем с натрием. Это позволяет использовать валиномицин как специфический калиевый ионофор. В липидной фазе молекула валиномицина имеет форму манжетки, устланной изнутри полярными группами, а снаружи – неполярными гидрофобными остатками молекул валина (рис. 44). Образуя комплекс с ионами калия, попадающими внутрь молекулы-манжетки, валиномицин приобретает лучшую растворимость в липидной фазе мембраны. Ионы калия удерживаются внутри молекулы за счет межзарядных взаимодействий. Молекулы валиномицина, оказавшиеся у поверхности мембраны, могут захватывать из окружающей среды ионы калия и переносить его через мембрану, возвращаясь обратно с каким-нибудь противоионом (например, протоном). Таким образом, происходит челночный перенос ионов калия через мембрану.

Рис. 44. Валиномицин ([L-лактатL-валин-D-оксиизовалериановая кислота-D-валин]3) – циклический депсипептидпереносчик иона калия/ Ион калия фиксируется в центре за счет межзарядных взаимодействий с участием карбонильных групп пептида (кружки).
Примером челночного переносчика является ионофор А23186, транспортирующий двухвалентные катионы Са 2+ по принципу ионного обмена: на каждый двухвалентный катион, переносимый в клетку, он «выносит» из клетки 2 Н+. В конечном итоге работа этого переносчика не приводит к деполяризации мембраны. Другими словами, ионофор А23187 осуществляет электронейтральный антипорт. Этот ионофор часто используется в экспериментах для повышения концентрации свободного кальция в цитозоле.
К образованию каналов в мембране способны антибиотики трех групп: грамицидины, аламетицин и полиеновые антибиотики. Общим для них является то, что их молекулы амфифильны, благодаря чему они способны встраиваться в мембрану и образовывать поры, пронизывая мембрану (в виде одной молекулы или димера). Заряженные или полярные группы размещены в таких молекулах на одном конце молекулы. Таким образом, благодаря внутри- и межмолекулярным связям полярных групп они способны погружаться в мембрану, образуя в ней каналы. Известны и другие ионофоры, способные переносить как одно-, так и двухвалентные ионы: – нактины, энниатины, макроциклические полиэфиры (крауны), жирорастворимые слабые кислоты (протонофоры) и др.
Как простая, так и облегченная диффузия (транспортные процессы, обеспечиваемые ионными каналами и переносчиками), осуществляется по градиенту концентрации или градиенту электрического заряда (объединяемым термином «электрохимический градиент»). Такие механизмы транспорта согласно классификации определяются как пассивный транспорт. Ионные каналы представляют собой сложные гликопротеиновые комплексы, осуществляющие быстрый пассивный транспорт ионов через биологические мембраны. Ионные каналы классифицируются по их проницаемости, селективности к различным ионам и по принципу открывания (закрывания) воротного механизма. Каналы способны избирательно взаимодействовать с определенными ионами (К + , Na + , Са 2+ , Сl – ) при изменении мембранного потенциала, гормональных, механических и осмотических воздействиях, которые через сенсор внешнего стимула влияют на работу воротного механизма канала.
Наиболее важными свойствами ионных каналов являются селективность, способность к инактивации и величина проводимости, а также фармакологическая характеристика, которые определяют кинетические свойства конкретного ионного канала. Зная проницаемость одиночного канала и суммарную проницаемость мембраны, можно рассчитать плотность ионных каналов. Например, плотность К + -каналов на плазмалемме достигает 5•10 11 каналов/м 2 , при этом они занимают 0,01% общей площади мембраны. Открывание и закрывание ионных каналов регулируется мембранным потенциалом, их ионным окружением (особенно ионами Са 2+ и рН), фосфорилированием протеинкиназ. Движение иона через пору канала сопряжено с преодолением энергетического барьера, величина которого зависит от диаметра, энергии гидратации иона, величины pH, ионной силы и других условий, способных изменять энергию активации при прохождении селективного фильтра.
В зависимости от способа управления воротным механизмом канала со стороны сенсора внешнего сигнала каналы делятся на две группы. Первую группу составляют такие типы каналов, у которых сенсор внешнего стимула непосредственно входит в состав молекулы канала. Эта группа включает в себя потенциал- и лиганд-зависимые каналы. Потенциал-зависимые ионные каналы (К + , Na + , Са 2+ , Сl – ) реагируют на изменение мембранного потенциала, лиганд-зависимые – открываются и закрываются при связывании с рецептором специфических агонистов, участвующих, например, в процессе быстрой передачи сигналов. У каналов второй группы сенсор внешнего стимула пространственно отделен от канала. В этом случае сигнал от сенсора передается на канал через систему внутриклеточных посредников. Эта группа включает в себя рецептор-зависимые каналы и каналы, управляемые G-белками (подробнее см. раздел о трансдукции).
Ионные каналы делятся на:
Все потенциал-зависимые каналы устроены, по-видимому, одинаково. Интегральный белок канальной структуры образует пору в мембране, кроме нее имеется селективный фильтр, обеспечивающий специфичность канала, а также сенсор мембранного потенциала – устройство, управляющее состоянием канала в зависимости от знака и величины потенциала. Известны каналы, обеспечивающие проводимость мембраны для Na + , K + , Ca 2+ , Na + – они активируются такими веществами, как грайанотоксином и вератридином. Действие этих соединений, как и эффект электрического потенциала, устраняется тетродотоксином. К + -каналы блокируются тетраэтиламмонием и его производными. Известны также Са-зависимые К + -каналы, открывающиеся при существенном возрастании концентрации внутриклеточного Са 2+ . Са 2+ -каналы также бывают потенциал-зависимыми и потенциал-индифферентными. Последние чувствительны к целому ряду разнообразных химических веществ, носящих название Са-антагонистов (верапамил, нифедипин и т.д.). Известны и агонисты Са-каналов. Открывание этих каналов осуществляется в присутствии адениновых нуклеотидов и следовых количеств кальция. Чувствительность Са-каналов к цАМФ и протеинкиназам объясняется тем, что фосфорилирование регуляторных компонентов канала контролирует длительность его нахождения в открытом состоянии.
Потенциал-зависимые каналы устроены таким образом, что интегральный белок канальной структуры образует пору в мембране. В канале выделяют внутреннее и наружное устья и пору, которая с помощью воротного механизма может открываться и закрываться. Гидрофильные аминокислоты выстилают стенки поры, а гидрофобные – контактируют с липидной фазой мембраны. В канале имеются селективный фильтр, обеспечивающий специфичность канала, и сенсор градиента электрического потенциала на мембране. Открывание и закрывание воротного механизма каналов является результатом конформационных изменений в белке. При открывании ионного канала регистрируется резкое возрастание электрического тока через мембрану.
Проницаемость каналов составляет от 10 6 до 10 8 ионов в секунду, что на три порядка выше, чем транспорт ионов, осуществляемый ионными насосами или переносчиками, и на 11 порядков выше, чем простая диффузия ионов через мембрану. Отличительной особенностью ионных каналов является то, что в открытом состоянии они обеспечивают относительно постоянный поток ионов в одном направлении при конкретном значении мембранного потенциала и в определенной ионной среде. Односторонняя проницаемость – еще одна особенность транспорта ионов через каналы. В процессе транспорта через канал происходит взаимодействие иона с белком, поэтому передвижение ионов по каналам отличается от их транспорта через водные поры, в которых эти взаимодействия сведены до минимума.
В противоположность вышеописанным процессам активный транспорт осуществляется против градиента концентрации или заряда, поэтому он требует притока дополнительной энергии, которая обычно обеспечивается за счет гидролиза АТФ (иногда – за счет мембранного потенциала). Первично-активный транспорт ионов в большинстве случаев осуществляется транспортными АТФазами (ионными насосами), источником энергии для которых является гидролиз АТФ или пирофосфата. В мембранах хлоропластов и митохондрий при работе систем первично-активного ионного транспорта источником энергии является трансмембранный потенциал, создаваемый работой окислительно-восстановительных цепей. Некоторые транспортные процессы осуществляются за счет гидролиза других макроэргических соединений, как, например, фосфоенолпирувата, или же за счет энергии света.
Первично-активный транспорт отличается от пассивного тем, что одна из его стадий является энергозависимой. Первично-активный транспорт ионов мембранными ферментами часто демонстрирует отклонения от кинетики Михаэлиса – Ментен, которые могут быть обусловлены свойствами самой системы переноса. Типичный пример первично-активного транспорта – активный транспорт ионов. Он осуществляется специальными ионными насосами, которые представляют собой интегральные белки клеточных мембран. Функция ионных насосов заключается в переносе ионов через мембрану против электрохимического градиента за счет энергии гидролиза АТФ. В соответствии с этим ионные насосы обладают аденозинтрифосфат-фосфогидролазной активностью. Последняя специфически контролируется переносимыми насосом ионами и требует для своей работы ионов магния. Таким образом, ион-транспортирующие системы являются Mg-АТФазами, приобретшими способность дополнительной регуляции переносимыми ионами как кофакторами. В процессе их функционирования энергия химической связи АТФ превращается в энергию электрохимического градиента переносимых этими системами ионов. По этой причине понятно, почему синонимом термина «ионные насосы» стал термин «транспортные АТФазы». Транспортные АТФазы классифицируют по переносимым ими (и активирующим их работу) ионам. Четыре типа АТФаз имеют отношение к транспорту ионов – протонная (К,Н-зависимая)-АТФаза, анионная (НСО 3-)-АТФаза, Са-АТФаза и Na/K-АТФаза.
При работе Nа/К–АТФазы за счет энергии высвобождающейся при гидролизе каждой молекулы АТФ в клетку переносятся два иона калия и выкачиваются три иона натрия. Таким образом, создается повышенная (по сравнению с межклеточной средой) концентрация в клетке ионов калия и пониженная концентрация ионов натрия, что имеет важное физиологическое значение. Са-насос обеспечивает активный перенос двух ионов кальция, а протонный насос – двух протонов на одну молекулу АТФ (рис. 45). У растений роль протонного насоса выполняет протонная пирофосфатаза (Н + -РРа).

Активный транспорт ионов может быть электронейтральным или электрогенным. Транспортная система электронейтральна, если при ее функционировании происходит обмен внутриклеточных ионов на внеклеточные в эквивалентных (по заряду) количествах. Если транспорт веществ через мембрану не сопровождается накоплением заряда, то такой транспорт называют электронейтральным.
Транспорт может быть электронейтральным при соблюдении следующих условий:
- при переносе незаряженного вещества;
- при симпорте катионов и анионов равной валентности;
- при антипорте двух равнозаряженных ионов.
В том случае, когда количество зарядов, переносимых за единицу времени в одном направлении, не компенсируется суммой зарядов, переносимых в противоположном направлении, транспортный механизм генерирует дополнительную разность потенциалов на мембране и называется электрогенным. Во время работы электрогенного насоса, например, Na/K-АТФазы, из клетки выносится 3 положительных заряда (3 иона Nа + ), а входят в клетку 2 положительных заряда (2 иона К + ), в результате чего в клетке происходит накопление положительных электрических зарядов, и на мембране возникает разность потенциалов. При работе электронейтрального насоса система активного транспорта является лишь средством поддержания концентрационных градиентов ионов и непосредственно не участвует в создании мембранного потенциала клетки. Существует классификация способов, которые обеспечивают транспорт веществ через клеточную мембрану (рис. 46).

Транспорт может идти по механизму унипорта (облегченной диффузии), согласно которому только одно вещество переносится через биомембрану в одном направлении с помощью канальных или транспортных белков (например, транспорт глюкозы в клетках печени). Примерами унипорта также являются транспорт ионов кальция через внутреннюю мембрану митохондрий. Транспортный процесс, который требует сопряженного переноса двух или более ионов (веществ) в одном направлении, называют симпортом, как, например, активный транспорт аминокислот или глюкозы вместе с ионами натрия в эпителиальных клетках кишечника протекает по механизму симпорта.
Транспорт ионов (веществ), сопряженный с переносом других ионов (веществ) в противоположном направлении, называют антипортом, как, например, обмен ионов НСО 3 – на Сl – в мембране эритроцитов. Примером такого транспорта может также служить Na + /H + антипорт во внутренней мембране митохондрий и K + /H + -антипорт, катализируемый ионофором нигерицином в бислоях. Трансмембранный обмен (антипорт) или однонаправленный транспорт (симпорт) рассматриваемых ионов осуществляют специфические переносчики.
Таким образом, при симпорте и антипорте белки функционируют в режиме, при котором перенос одного растворенного вещества зависит от одновременного или последовательного переноса другого вещества. Указанные термины – антипорт и симпорт (котранспорт) – связывают не только с явлениями пассивного транспорта, осуществляющегося с помощью переносчика (например, симпорт анионов, осуществляющийся методом электрофореза), но и в случае, когда процесс сопряжен с энергозависимыми реакциями.
Энергия, запасаемая клеткой в виде ионных градиентов, созданных первично активным транспортом веществ через мембрану, может использоваться другими транспортными процессами, которые в таком случае (по отношению к энергетическому обеспечению процесса) называют вторично–активным транспортом. Так, к примеру, происходит при аккумуляции в клетке глюкозы или других метаболитов – они переносятся против концентрационного градиента за счет того, что одновременно с ними в клетку переносятся ионы Na + . В клетках низших эукариот градиент Na + может быть заменен градиентом протонов. Перенос осуществляется с помощью белков-переносчиков, имеющих молекулярную массу 25– 40 кДа. Некоторые переносчики сахаров выделены из плазматических мембран слизистой кишечника, эритроцитов, бактерий. Они имеют широко варьирующее сродство (K а для переносимых веществ варьирует от 10 -7 до 10 -4 М) и различную специфичность к переносимым соединениям. Обнаруживаются случаи переноса одной и той же аминокислоты 3–4 разными переносчиками или, наоборот, выявляется специфичность одного переносчика к 2–3 разным соединениям. Удобным объектом для изучения транспорта метаболитов являются микроорганизмы, дефицитные по конкретным переносчикам. Соответствующие субстраты не попадают внутрь клетки и поэтому не могут утилизироваться.
К вторично-активному транспорту относятся и процессы переноса, сопряженные с ферментативной модификацией переносимых соединений. Например, фосфотрансферазная система бактерий, отсутствующая у эукариот, фосфорилирует сахара в процессе их проникновения через мембрану, вовлекая их тем самым в клеточный метаболизм. У грам-отрицательных бактерий так переносятся D-глюкоза, D-фруктоза и D-глюкозамин. У грам- положительных бактерий набор переносимых веществ шире: сюда относятся также пентозы, сахароза, трегалоза, лактоза, глицерин. При этом лактоза и фруктоза фосфорилируются по С 1, остальные вещества – по концевому углероду.
Митохондрии – это пример системы, осуществляющей вторично-активный транспорт Са 2+ . В этом случае перенос иона осуществляется по градиенту потенциала, образованного на мембране за счет метаболических процессов, например, дыхания или гидролиза АТФ. При этом на мембране генерируется потенциал, и транспорт Са 2+ осуществляется по принципу электрофореза.
http://poisk-ru.ru/s53930t4.html
http://farmf.ru/lekcii/harakteristika-transportnyh-protsessov-v-kletochnoj-membrane/