При аэробном дыхании на каждую окисленную молекулу глюкозы образуется 38 молекул АТФ.
Общее количество энергии, высвобождаемой при полном окислении глюкозы, составляет 2880 кДж на 1 моль.
В одном моле АТФ заключено 30,6 кДж. В 38 молях АТФ заключено 30,6 х38 = 1162,8 кДж.
Таким образом, эффективность превращения энергии при аэробном дыхании составляет: 1162,8/2880 = 40,4%.
Анаэробное дыхание
1. ДРОЖЖИ (СПИРТОВОЕ БРОЖЕНИЕ). При спиртовом брожении на каждую молекулу глюкозы образуются две молекулы АТФ.
Общее количество энергии, высвобождаемой из глюкозы при ее превращении в этанол, составляет 210 кДж на 1 моль.
В двух молях АТФ заключено 2 х30,6 = 61,2 кДж.
Следовательно, эффективность превращения энергии при спиртовом брожении составляет 61,2/210 = 29,1%.
2. МЫШЦЫ (МОЛОЧНОКИСЛОЕ БРОЖЕНИЕ). При молочнокислом брожении на каждую молекулу глюкозы образуются две молекулы АТФ.
Глюкоза————> 2Молочная кислота + 2АТФ
Общее количество энергии, высвобождаемой из глюкозы при ее превращении в молочную кислоту, составляет 150 кДж на 1 моль.
Таким образом, эффективность превращения энергии при молочнокислом брожении составляет 61,2/150 = 40,8%.
Приведенные цифры показывают, что эффективность превращения энергии в каждой из этих систем довольно высокая по сравнению с бензиновым (25-30%) или паровым (8-12%) двигателями. Количество же энергии, запасаемое в виде АТФ при аэробном дыхании, в 19 раз больше, чем при анаэробном (38 молекул АТФ на одну молекулу глюкозы в первом случае и 2 молекулы АТФ — во втором). С этой точки зрения аэробное дыхание значительно эффективнее анаэробного. Связано это с тем, что при анаэробном дыхании значительная часть энергии остается «запертой» в этаноле или молочной кислоте. Энергия, заключенная в этаноле, так и остается для дрожжей навсегда недоступной и, значит, спиртовое брожение в смысле получения энергии — малоэффективный процесс. Из молочной же кислоты позднее может быть извлечено довольно большое количество энергии, если появится кислород. В присутствии кислорода молочная кислота превращается в печени в пировиноградную кислоту. Последняя поступает затем в цикл Кребса и полностью окисляется до С02 и Н2О, в результате чего дополнительно образуется большое количество молекул АТФ. Возможен и другой путь — за счет энергии АТФ из пировиноградной кислоты может вновь образоваться глюкоза в процессе, который представляет собой обращение гликолиза.
Анаэробное и аэробное дыхание
Процесс дыхания состоит из двух основных этапов. Первый, начальный, — анаэробное дыхание, в результате которого дыхательный субстрат (углеводы) распадается до простейших продуктов типа пировиноградной кислоты. Дальше превращение пировиноградной кислоты может проходить двумя путями: кислородным до конечных продуктов СО2 и Н2О или анаэробным по типу брожения. Таким образом, устанавливается определенная связь между дыханием и брожением. Учение о генетической связи между этими процессами было разработано С. П. Костычевым. Общую схему ставить в таком виде:
Л. Пастер первым доказал, что высшие растения не прекращают выделения углекислого газа и после того, как попадают в среду, лишенную кислорода. Но дыхание растений в этих условиях сопровождается накоплением в их тканях спирта. Дыхание за счет связанного кислорода называют также интрамолекулярным. Его можно представить уравнением $$\large \ceO_6 = 2C_2H_5OH + 2CO_2 +117 Дж>$$
При дыхании такого типа жизнь зеленого растения продолжается недолго — оно погибает. Очевидно, происходит отравление продуктами обмена, которые образуются в этом случае.
Низкий энергетический эффект брожения также имеет существенное значение. Спирт содержит большой запас энергии, которая не используется при интрамолекулярном дыхании. Установлено, что для получения того же количества энергии в анаэробных условиях ткани высшего растения должны вследствие низкого энергетического уровня этого процесса израсходовать в 30-50 раз больше пластических веществ, чем при аэробном дыхании. В результате анаэробного дыхания ткани истощаются — они лишаются различных промежуточных веществ, которые обычно образуются при кислородном дыхании.
Существует взгляд на интрамолекулярное дыхание (брожение) у высших растений как на рудиментарную функцию. Наряду с аэробным дыханием в тканях растений в той или иной мере всегда происходят и процессы брожения. В тканях растений, нормально снабжающихся кислородом, они обнаружены во многих случаях. Так, продукты спиртового брожения (уксусный альдегид, этиловый спирт) накапливаются в интенсивно растущих органах растений, в сочных тканях различных плодов — лимонов, яблок, мандаринов.
Брожение, и дыхание тесно связаны между собой. Об этом свидетельствует прежде всего тот факт, что в растении найдены те же промежуточные продукты, которые образуются в дрожжах при спиртовом брожении. Так, во многих растениях обнаружены глюкозо-6-фосфат, фруктозо-6-фосфат, фруктозо-1,6-дифосфат. Эти фосфорные эфиры сахаров оказались в листьях гороха, сахарной свеклы, овса, ячменя, в прорастающих семенах гороха. В листьях ячменя обнаружены фосфоглицериновая и пировиноградная кислоты, в луке — пировиноградная кислота, в некоторых плодах — уксусный альдегид. Все эти соединения являются промежуточными продуктами спиртового брожения. О единстве и теснейшей связи процессов брожения и дыхания свидетельствует также то, что в растениях выявлены ферменты, катализирующие спиртовое брожение.
При аэробном дыхании последним акцептором водорода является кислород, и поэтому конечным продуктом окисления оказывается вода. При анаэробном дыхании к последним акцепторам водорода относятся другие вещества, образующиеся в процессе брожения субстрата. В. И. Палладии еще в 1912 г. указывал, что при аэробном дыхании весь водород глюкозы окисляется до воды исключительно кислородом воздуха. Образование спирта при брожении возможно потому, что последним акцептором водорода в этом случае является альдегид.
Экспериментально доказано, что дегидрогеназы и их коферменты НАД и НАДФ, катализирующие отщепление водорода, от окисляемых органических веществ, играют важную роль в процессах тканевого дыхания и при разных видах анаэробных брожений (спиртовом, молочнокислом, маслянокислом и др.).
Восстановленная форма никотинамидадениндииуклеотидфосфата (НАДФН2) может отдавать водород таким его ацепторам, как пировиноградная кислота: $$\large \ce>\underset<\text<молочная кислота>><\ce>$$
В других случаях восстановленный НАДФ может отдавать свой водород ацетальдегиду: $$\large \ce>\underset<\text<этиловый спирт>><\ce>\; + НАДФ$$
Таким образом, в зависимости от того, к какому акцептору будет присоединен с помощью дегидрогеназ водород, образуются и соответствующие продукты, определяющие тип брожения (молочнокислое, спиртовое и др.). Для взаимодействия между акцептором водорода и соответствующей кодегидразой необходимо присутствие в клетке соответствующего фермента. Следовательно, анаэробное окисление не является патологическим процессом для высших растений. Наряду с аэробным дыханием брожение, очевидно, — один из постоянных процессов окислительного газообмена в их тканях. В разных тканях при различных условиях участие брожения в дыхательном газообмене может значительно изменяться. Но при этом анаэробные процессы в зависимости от внутренних и внешних условий происходят, очевидно, всегда.
Химизм анаэробной фазы дыхания (гликолиз)
Начальный этап анаэробного распада углеводов заключается в образовании ряда фосфорных эфиров сахаров (гексоз). Важную роль фосфорной кислоты в процессе анаэробного дыхания впервые установили русские биохимики Л. А. Иванов и А. Н. Лебедев, которые экспериментально доказали образование в этом процессе соединений сахара с фосфорной кислотой. С помощью различных ингибиторов было выяснено, что анаэробному распаду при брожении подвергается не свободная молекула гексозы, а ее фосфорный эфир, который образовался из гексозы и фосфорной кислоты и является активным и лабильным соединением в отличие от химически инертной молекулы гексозы. Активация молекулы гексозы, повышение ее реакционной способности происходят постепенно и проходят ряд этапов.
На первом этапе брожения и дыхания молекула глюкозы под действием фермента гексокииазы принимает остаток фосфорной кислоты от АТФ, которая превращается в АДФ, и в результате образуется глюкопиранозо-6-фосфат, превращающийся под действием фермента фосфогексоизомеразы (оксоизомеразы) в фруктофуранозо-6-фосфат. На дальнейшем этапе гликолиза фрукто- фуранозо-6-фосфата к нему присоединяются еще один остаток фосфорной кислоты. Источником энергии для образования данного эфира является также молекула АТФ. Эту реакцию катализирует фосфогексокиназа, активируемая ионами магния. В результате образуются фруктофуранозо-1,6-дифосфат и новая молекула аденозиндифосфата:
Далее молекула фруктозо-1,6-дифосфата под влиянием фермента альдолазы расщепляется на две молекулы фосфотриоз: фосфодиоксиацетон и 3-фосфоглицериновый альдегид. Фоcфодиоксиацетон под действием фермента фосфотриозоизомеразы полностью превращается в 3-фосфоглицериновый альдегид.
Следующий этап гликолиза заключается в окислении 3-фосфоглицеринового альдегида специфической дегидрогеназой и фосфорилировании глицериновой кислоты с использованием минеральной фосфорной кислоты. Образовавшаяся в результате этой реакции 1,3-дифосфоглицериновая кислота передает при участии фермента фосфоферазы один остаток фосфорной кислоты молекуле АДФ, которая превращается в АТФ, при этом образуется 3-фосфоглицериновая кислота. Последняя под действием фермента фосфоглицеромутазы переходит в 2-фосфоглицериновую кислоту, которая под влиянием фермента енолазы превращается в фосфоенолпировиноградную кислоту и наконец в пировиноградную кислоту. Процесс преобразования 3-фосфоглицеринового альдегида в 1,3-дифосфоглицериновую кислоту осуществляется по схеме:
Процесс превращения 1,3-дифосфоглицернновой кислоты пировиноградную происходит по такой схеме:
На рисунке выше приведены общая схема реакций распада глюкозы до пировиноградной кислоты в анаэробной фазе дыхания и обратные реакции, в результате которых из пировиноградной кислоты синтезируется глюкоза. Ферменты гликолитического распада глюкозы легко экстрагируются из клеток, поэтому считают, что они локализуются в растворимой части цитоплазмы.
Образованием пировиноградной кислоты из фосфоенолпирувата заканчивается гликолитическое расщепление гексозы. На каждый моль использованной в этих реакциях гексозы расходуются два моля АТФ, тогда как в реакциях превращения двух молекул 1,3-дифосфоглицериновой кислоты и двух молекул фосфоенолпировиноградной кислоты синтезируются четыре молекулы АТФ, в результате остаются неиспользованными две молекулы АТФ. Кроме того, в ходе окисления гексозы до пирувата восстанавливаются две молекулы НАДН2 или НАДФН2 (в зависимости от растений); каждая из них, окисляясь, образует по три молекулы АТФ, а всего — шесть молекул АТФ. Таким образом, при гликолитическом распаде гексозы, который является начальным этапом анаэробного дыхания, происходит потребление двух молекул АТФ, регенерация АДФ и синтез четырех новых молекул аденозинтрифосфата. Пировиноградная кислота, образовавшаяся в результате описанных реакций в анаэробных условиях, подвергается превращениям, которые осуществляются при спиртовом или молочнокислом брожении.
Химизм аэробной фазы дыхания
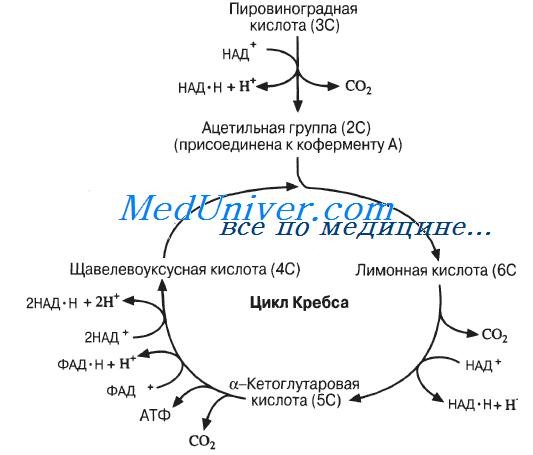
В аэробных условиях пировиноградная кислота в растениях окисляется полностью до СО2 и Н2О. Это окисление, как установлено английским биохимиком Г. Кребсом, проходит последовательно с образованием ди- и трикарбоновых кислот, поэтому оно называется циклом ди- и трикарбоновых кислот, или лимоннокислым, или циклом Кребса, которым завершается окисление продуктов распада углеводов, жиров и белков. В результате, молекула пировиноградной кислоты полностью окисляется до. трех молекул углекислого газа и двух молекул воды: CH3COCOOH+2½O2 → 3CO2+2H2O $$\large \ce 3CO_2 + 2H_2O>$$
Установлено, что в процессе постепенного, окисления пировиноградной кислоты образуются различные промежуточные органические кислоты с четырьмя или шестью атомами углерода, встречающиеся в растениях.
Предложенная Г. Кребсом схема является дальнейшим развитием учения С. П. Костычева о генетической связи дыхания и брожения.
Рис. 41. Схема реакций анаэробного распада углеводов
Рис. 42. Цикл ди- и трикарбоновых кислот (цикл Кребса)
Таким образом, цикл Кребса заключается в образовании лимонной кислоты из щавелевоуксусной кислоты и ацетилкоэнзима А (ацетил-КоА) и регенерации щавелевоуксусной кислоты из лимонной.
Первая реакция в цикле — образование промежуточного продукта «активированной» уксусной кислоты в виде ацетил-КоА, который окончательно окисляется. Энергия, выделяющаяся в пяти реакциях окисления, фиксируется в виде макроэргических пирофосфатных связей аденозинтрифосфата. Это — следующие окислительно-восстановительные реакции: образование ацетил-коэнзима А; окисление лимонной и изолимонной кислот через цис-аконитовую до щавелевоянтарной кислоты, α-кетоглутаровой — до сукцинил-КоА, янтарной — до фумаровой, яблочной — до щавелевоуксусной, которая является основным соединением в цикле: она катализирует полный распад пировиноградной кислоты, после чего происходит регенерация щавелевоуксусной кислоты в циклическом процессе.
Таким образом, с каждым оборотом цикла исчезает одна молекула пировиноградной кислоты и от различных компонентов цикла отщепляется три молекулы СО2 и пять пар атомов водорода (электронов).
Органические кислоты, входящие в цикл, имеются в тканях почти всех растений. В растениях найдены также все важнейшие ферментные системы, участвующие в превращении этих органических кислот, а именно: аконитаза, дегидрогеназы изолимонной, яблочной и янтарной кислот, фумараза, карбоксилаза щавелевоянтарной кислоты. Все ферменты цикла трикарбоновых кислот сконцентрированы в матриксе митохондрий; здесь также обнаружены ферменты окисления жирных кислот и др. Считают, что основное назначение цикла. Кребса заключается в подготовке материала для синтетических процессов, происходящих во время роста молодых клеток. На такие процессы расходуются промежуточные продукты цикла: α-кетоглутаровая, фумаровая кислоты и др. Эти соединения могут быть исходными веществами для многочисленных реакций синтеза и обмена аминокислот, синтеза нуклеотидов, образования различных циклических соединений, жиров и других веществ.
Дополнительные материалы по теме:
Аэробное и анаэробное дыхание растений
1. Аэробное дыхание
1.1 Окислительное фосфолирование
2. Анаэробное дыхание
2.1 Типы анаэробного дыхания
Дыхание присуще всем живым организмам. Оно представляет собой окислительный распад органических веществ, синтезированных в процессе фотосинтеза, протекающих с потреблением кислорода и выделением диоксида углерода. А.С. Фаминцын рассматривал фотосинтез и дыхание как две последовательные фазы питания растений: фотосинтез готовит углеводы, дыхание перерабатывает их в структурную биомассу растения, образуя в процессе ступенчатого окисления реакционноспособные вещества и освобождая энергию, необходимую для их превращения и процессов жизнедеятельности в целом. Суммарное уравнение дыхания имеет вид:
C H O + 6O → 6CO + 6H O + 2875кДж.
Из этого уравнения становится ясно, почему именно скорость газообмена используют для оценки интенсивности дыхания. Оно было предложено в 1912 г. В. И. Палладиным, который считал, что дыхание состоит из двух фаз – анаэробной и аэробной. На анаэробном этапе дыхания, идущем в отсутствие кислорода, глюкоза окисляется за счет отнятия водорода (дегидрирования), который, по мнению ученого, передается на дыхательный фермент. Последний при этом восстанавливается. На аэробном этапе происходит регенерация дыхательного фермента в окислительную форму. В. И. Палладин впервые показал, что окисление сахара идет за счет непосредственного окисления его кислородом воздуха, поскольку кислород не встречается с углеродом дыхательного субстрата, а связано с его дегидрированием.
Существенный вклад в изучение сути окислительных процессов и химизма процесса дыхания внесли как отечественные (И.П. Бородин, А.Н.Бах, С.П. Костычев, В.И. Палладин), так и зарубежные (А.Л. Лавуазье, Г. Виланд, Г. Кребс) исследователи.
Жизнь любого организма неразрывно связана с непрерывным использованием свободной энергии, генерируемой при дыхании. Неудивительно, что изучению роли дыхания в жизни растения в последнее время отводят центральное место в физиологии растений.
1. Аэробное дыхание
Аэробное дыхание – это окислительный процесс, в ходе которого расходуется кислород. При дыхании субстрат без остатка расщепляется до бедных энергией неорганических веществ с высоким выходом энергии. Важнейшими субстратами для дыхания служат углеводы. Кроме того, при дыхании могут расходоваться жиры и белки.
Аэробное дыхание включает два основных этапа:
— бескислородный, в процессе, которого происходит постепенное расщепление субстрата с высвобождением атомов водорода и связыванием с коферментами (переносчиками типа НАД и ФАД);
— кислородный, в ходе которого происходит дальнейшее отщепление атомов водорода от производных дыхательного субстрата и постепенное окисление атомов водорода в результате переноса их электронов на кислород.
На первом этапе вначале высокомолекулярные органические вещества (полисахариды, липиды, белки, нуклеиновые кислоты и др.) под действием ферментов расщепляются на более простые соединения (глюкозу, высшие карбоновые кислоты, глицерол, аминокислоты, нуклеотиды и т.п.) Этот процесс происходит в цитоплазме клеток и сопровождается выделением небольшого количества энергии, которая рассеивается в виде тепла. Далее происходит ферментативное расщепление простых органических соединений.
Примером такого процесса является гликолиз – многоступенчатое бескислородное расщепление глюкозы. В реакциях гликолиза шестиуглеродная молекула глюкозы (С ) расщепляется на две трехуглеродные молекулы пировиноградной кислоты (С ). При этом образуется две молекулы АТФ, и выделяются атомы водорода. Последние присоединяются к переносчику НАД (никотинамидадениндинклеотид), который переходит в свою восстановительную форму НАД ∙ Н + Н . НАД кофермент, близкий по своей структуре к НАДФ. Оба они представляют собой производные никотиновой кислоты – одного из витаминов группы В. Молекулы обоих коферментов электроположительны (у них отсутствует один электрон) и могут играть роль переносчика как электронов, так и атомов водорода. Когда акцептируется пара атомов водорода, один из атомов диссоциирует на протон и электрон:
Н → Н + е ,
а второй присоединяется к НАД или НАДФ целиком:
НАД + Н + [Н + е ] → НАД ∙ Н + Н.
Свободный протон позднее используется для обратного окисления кофермента. Суммарно реакция гликолиза имеет вид
C H O +2АДФ + 2Н РО + 2 НАД →
2С Н О + 2АТФ + 2 НАД ∙ Н + Н + 2 HO
Продукт гликолиза – пировиноградная кислота (С Н О ) – заключает в себе значительную часть энергии, и дальнейшее ее высвобождение осуществляется в митохондриях. Здесь происходит полное окисление пировиноградной кислоты до CO и H O. Этот процесс можно разделить на три основные стадии:
На первой стадии пировиноградная кислота взаимодействует с веществом, которое называют коферментом А, в результате чего образуется ацетилкофермент а с высокоэнергетической связью. При этом от молекулы пировиноградной кислоты отщепляется молекула CO (первая) и атомы водорода, которые запасаются в форме НАД ∙ Н + Н .
Вторая стадия – цикл Кребса (рис. 1)
В цикл Кребса вступает ацетил–КоА, образованный на предыдущей стадии. Ацетил–КоА взаимодействует со щавелево-уксусной кислотой, в результате образуется шестиуглеродная лимонная кислота. Для этой реакции требуется энергия; ее поставляет высокоэнергетическая связь ацетил–КоА. В конце цикла щавелево-лимонная кислота регенерируется в прежнем виде. Теперь она способна вступить в реакцию с новой молекулой ацетил–КоА, и цикл повторяется. Суммарно реакция цикла может быть выражена следующим уравнением:
ацетил-КоА + 3H O + 3НАД + ФАД + АДФ + Н РО →
КоА + 2CO + 3НАД ∙ Н + Н +ФАД ∙ H + АТФ.
Таким образом, в результате распада одной молекулы пировиноградной кислоты в аэробной фазе (декарбоксилирование ПВК и цикла Кребса) выделяется 3CO , 4 НАД ∙ Н + Н , ФАД ∙ H . Суммарно реакцию гликолиза, окислительного декарбоксилирования и цикла Кребса можно записать в следующем виде:
C H O + 6 H O + 10 НАД + 2ФАД →
6CO + 4АТФ + 10 НАД ∙ Н + Н + 2ФАД ∙ H .
Третья стадия – электротранспортная цепь.
Пары водородных атомов, отщепляемые от промежуточных продуктов в реакциях дегидрирования при гликолизе и в цикле Кребса, в конце концов, окисляются молекулярным кислородом до H O с одновременным фосфолированием АДФ в АТФ. Происходит это тогда, когда водород, отделившийся от НАД ∙ H и ФАД ∙ H , передается по цепи переносчиков, встроенных во внутреннюю мембрану митохондрий. Пары атомов водорода 2Н можно рассматривать как 2 Н + 2е . Движущей силой транспорта атомов водорода в дыхательной цепи является разность потенциалов.
С помощью переносчиков ионы водорода Н переносятся с внутренней стороны мембраны на ее внешнюю сторону, иначе говоря, из матрикса митохондрии в межмембранное пространство (рис. 2).
При переносе пары электронов от над на кислород они пересекают мембрану три раза, и этот процесс сопровождается выделением на внешнюю сторону мембраны шести протонов. На заключительном этапе протоны переносятся на внутреннюю сторону мембраны и акцептируются кислородом:
½ O + 2е → O .
В результате такого переноса ионов Н на внешнюю сторону мембраны митохондрий в перимитохондриальном пространстве создается концентрация их, т.е. возникает электрохимический градиент протонов .
Когда протонный градиент достигает определенной величины, ионы водорода из Н -резервуара движутся по специальным каналам в мембране, и их запас энергии используется для синтеза АТФ. В матриксе они соединяются с заряженными частичками О , и образуется вода: 2Н + О²ˉ → H O.
1.1 Окислительное фосфолирование
Процесс образования АТФ в результате переноса ионов Н через мембрану митохондрии получил название окислительного фосфолирования. Он осуществляется при участии фермента АТФ-синтетазы. Молекулы АТФ-синтетазы располагаются в виде сферических гранул на внутренней стороне внутренней мембраны митохондрий.
В результате расщепления двух молекул пировиноградной кислоты и переноса ионов водорода через мембрану по специальным каналам синтезируется в целом 36 молекул АТФ (2 молекулы в цикле Кребса и 34 молекулы в результате переноса ионов Н через мембрану).
Суммарное уравнение аэробного дыхания можно выразить следующим образом:
C H O + O + 6H O + 38АДФ + 38Н РО→
6CO + 12H O + 38АТФ
Совершенно очевидно, что аэробное дыхание прекратится в отсутствии кислорода, поскольку именно кислород служит конечным акцептором водорода. Если клетки не получают достаточного количества кислорода, все переносчики водорода вскоре полностью насытятся и не смогут передавать его дальше. В результате основной источник энергии дл образования АТФ окажется блокированным.
аэробное дыхание окисление фотосинтез
2. Анаэробное дыхание
Анаэробное дыхание. Некоторые микроорганизмы способны использовать для окисления органических или неорганических веществ не молекулярный кислород, а другие окисленные соединения, например, соли азотной, серной и угольной кислот, превращающиеся при этом в более восстановленные соединения. Процессы идут в анаэробных условиях, и их называют анаэробным дыханием:
2HNO + 12Н → N + 6H O + 2Н
H SO + 8Н → H S + 4HO
У микроорганизмов, осуществляющих такое дыхание, конечным акцептором электронов будет не кислород а неорганическое соединения – нитриты, сульфаты и карбонаты. Таким образом, различия между аэробным и анаэробным дыханием заключается в природе конечного акцептора электронов.
2.1 Типы анаэробного дыхания
Основные типы анаэробного дыхания приведены в таблице 1. есть также данные об использовании бактериями в качестве акцепторов электронов Mn , хроматов, хинонов и др.
Таблица 1 Типы анаэробного дыхания у прокариот (по: М.В Гусев, Л.А. Минеева 1992, с изменениями)
Конечный акцептор электронов
Нитратное дыхание и нитрификация
NO , NO
NO , NO, N O, N
Сульфатное и серное дыхание
SO , S
H S
Fe
Fe
CO
СН , ацетат
Свойство организмов переносить электроны на нитраты, сульфаты и карбонаты обеспечивает в достаточной степени полное окисление органического или неорганического вещества без использования молекулярного кислорода и обуславливает возможность получения большого количества энергии, чем при брожении. При анаэробном дыхании выход энергии только на 10% ниже. Чем при аэробном. Организмы, для которых характерно анаэробное дыхание, имеют набор ферментов электронтранспортной цепи. Но цитохромоксилаза в них заменяется нитратредуктазой (при использовании в качестве акцептора электронов нитрата) или аденилсульфатредуктазой (при использовании сульфата) или другими ферментами.
Организмы, способные осуществлять анаэробное дыхание за счет нитратов, — факультативные анаэробы. Организмы, использующие сульфаты в анаэробном дыхании, относятся к анаэробам.
Органические вещества из не органических зеленое растение образует только на свету. Эти вещества используются растением только для питания. Но растения не только питаются. Они дышат, как все живые существа. Дыхание происходит непрерывно днем о ночью. Дышат все органы растения. Растения дышат кислородом, а выделяют углекислый газ, как животные и человек.
Дыхание растений может происходить, как в темноте, так и на свету. Значит, на свету в растении протекают два противоположных процесса. Один процесс — фотосинтез, другой – дыхание. Во время фотосинтеза создаются органические вещества из неорганических и поглощается энергия солнечного света. Во время дыхания в растении расходуются органические вещества. А энергия, необходима для жизнедеятельности, освобождается. На свету в процессе фотосинтеза растения поглощают углекислый газ и выделяют кислород. Вместе с углекислым газом растения на свету поглощают из окружающего воздуха и кислород, необходимый растениям для дыхания, но в гораздо меньших количествах, чем выделяются при образовании сахара. Углекислого газа при фотосинтезе растения поглощают гораздо больше, чем выделяют его придыхании. Декоративные растения в комнате при хорошем освещении выделяют днем значительно больше кислорода, чем поглощают его в темноте ночью.
Дыхание во всех живых органов растения происходит непрерывно. Когда прекращается дыхание, растение, так же как и животное погибает.
1. Физиология и биохимия сельскохозяйственных растений Ф50/Н.Н. Третьяков, Е.И. Кошкин, Н.М. Макрушин и др.; под. ред. Н.Н. Третьякова. – М.; Колос, 2000 – 640 с.
2. Биология в экзаменационных вопросах и ответах Л44/ Лемеза Н.А., Камлюк Л.В.; 7-е изд. – М.: Айрис-пресс, 2003. – 512 с.








 H
H  O
O  → 6CO
→ 6CO  ) расщепляется на две трехуглеродные молекулы пировиноградной кислоты (С
) расщепляется на две трехуглеродные молекулы пировиноградной кислоты (С  ). При этом образуется две молекулы АТФ, и выделяются атомы водорода. Последние присоединяются к переносчику НАД
). При этом образуется две молекулы АТФ, и выделяются атомы водорода. Последние присоединяются к переносчику НАД  (никотинамидадениндинклеотид), который переходит в свою восстановительную форму НАД ∙ Н + Н
(никотинамидадениндинклеотид), который переходит в свою восстановительную форму НАД ∙ Н + Н  + е
+ е  ,
, РО
РО  + 2 НАД
+ 2 НАД 
 O + 3НАД
O + 3НАД  + ФАД + АДФ + Н
+ ФАД + АДФ + Н  РО
РО  →
→ O
O  . Движущей силой транспорта атомов водорода в дыхательной цепи является разность потенциалов.
. Движущей силой транспорта атомов водорода в дыхательной цепи является разность потенциалов.
 → O
→ O  , и образуется вода: 2Н
, и образуется вода: 2Н  O.
O. через мембрану).
через мембрану). H
H  →
→ + 12Н
+ 12Н  → N
→ N  , хроматов, хинонов и др.
, хроматов, хинонов и др. , NO
, NO 
 O, N
O, N , S
, S 

