Процесс гликолиза его реакции, аэробный и анаэробный (Таблица, схема)
Гликолиз — процесс окисления глюкозы, при котором из одной молекулы глюкозы образуются две молекулы пировиноградной кислоты, не является мембранозависимым процессом. Он происходит в цитоплазме. Однако ферменты гликолиза связаны со структурами цитоскелета. Суть гликолиза состоит в том, что молекула глюкозы (C6H12O6) без участия кислорода распадается на две молекулы пировиноградной кислоты (СН3СОСООН). При этом окисление идет за счет отщепления от молекулы глюкозы четырех атомов водорода, связывающихся со сложным органическим веществом НАД с получением двух молекул НАД•Н. Выделяющаяся при этом энергия запасается (40% от общего количества) в виде макроэргических связей двух молекул АТФ. 60% энергии выделяется в виде тепла. При последующем окислении НАД•Н получается еще 6 молекул АТФ. Таким образом, полный энергетический выход гликолиза в анаэробных условиях составляет 8 молекул АТФ.
Аэробный процесс гликолиза (10 реакций), уравнение (с образованием пирувата):
Анаэробный процесс гликолиза (11 реакций), уравнение (с образованием лактата):
Схема процесса гликолиза и его реакции

На схеме в рамках обозначены исходные субстраты и конечные продукты гликолиза, цифрами в скобках — число молекул.
ATP (АТФ) — это аденозинтрифосфорная кислота, универсальный источник энергии
ADP (АДФ) — это аденозиндифосфат, нуклеотид, участвует в энергетическом обмене
NAD (НАД) — никотинамидадениндинуклеотидфосфата
NADH (НАД•Н) — востановленная форма NAD
Таблица процесс гликолиза его реакции
Для распада и частичного окисления молекулы глюкозы требуется протекание 11 сложных последовательных реакций.
Ферменты, Активаторы, ингибиторы
Подготовительная стадия гликолиза
Стадия активации глюкозы проходит в 5 реакций, в ходе которых 1 молекула гексозы (глюкозы) расщепляется на 2 молекулы триоз-глицеральдегидфосфата
1. Необратимая реакция фосфорилирования глюкозы
Процесс гликолиза начинается с фосфорилирования глюкозы за счет АТФ — первая реакция. Это первая пусковая реакция гликолиза. Ее результатом является глюкозо-6-фосфат, имеющий отрицательный заряд. В гликолизе может участвовать не только глюкоза, но и другие гексозы (фруктоза), но в результате фосфорилирования и активации все равно образуется глюкозо-6-фосфат.

Ингибиторы: глюкозо-6-Ф, фосфоенолпируват.
2. Обратимая реакция изомеризации глюкозо-6-фосфата
Во второй реакции происходит изомеризация (внутримолекулярные перестройки) глюкозо-6-фосфата во фруктозо-6-фосфат.

3. Необратимая реакция фосфорилирования фруктозо-6-фосфата (ключевая стадия гликолиза)
В третьей реакции происходит фосфорилирование (присоединение остатка ортофосфорной кислоты) фруктозо-6-фосфата с образованием фруктозо-1,6-дифосфата. При этом затрачивается еще одна молекула АТФ (уже вторая) — это вторая пусковая реакция гликолиза. Она идет в присутствии Mg2 + и является необратимой, так как сопровождается масштабным уменьшением свободной энергии.

Ингибиторы: АТФ, цитрат, НАДН.
4. Обратимая реакция дихотомического расщепления фруктозо-1,6-дифосфата
В четвертой реакции гликолиза происходит расщепление фруктозо-1,6-дифосфата на две молекулы глицеральдегид-3-фосфата.

5. Обратимая реакция изомеризации дигидроксиацетона-3-фосфат в глицеральдегид-3-фосфат
В пятой реакции происходит изомеризация полученных триозофосфатов. На этом заканчивается первая стадия гликолиза.

Стадия генерации АТФ
Проходит в 6 реакций (или 5), в ходе которых энергия окислительных реакций трансформируется в химическую энергию АТФ (субстратное фосфорилирование).
6. Окисление глицеральдегид-3-фосфата до 1,3-дифосфоглицерата (реакция гликолитической оксиредукции)
В шестой реакции происходит окисление альдегидной группы до карбоксильной. Выделившийся Н+ акцептируется NAD, который восстанавливается до NADH. Освобождающаяся энергия затрачивается для образования высокоэнергетической связи 1,3-бифосфоглицерата (1,3-бифосфоглицериновая кислота).

7. Субстратное фосфорилирование АДФ (7)
В седьмой реакции фосфорильная группа 1,3-бифосфоглицерата переносится на ADP, в результате чего образуется АТР (напоминаем, что следует иметь в виду две параллельные цепи реакций, с участием двух молекул триоз, образовавшихся из одной молекулы гексозы, следовательно, синтезируется не одна, а две молекулы АТР).

8. Реакция изомеризации 3-фосфоглицерата в 2-фосфоглицерат
В восьмой реакции гликолиза происходит перенос фосфатной группы с третьего атома углерода на второй. В результате образуется 2-фосфоглицерат (2-фосфоглицериновая кислота).

9. Реакция енолизации
Девятая реакция сопровождается внутримолекулярными окислительно-восстановительными процессами, в результате которых образуется фосфоенолпируват (фосфоенолпировиноградная кислота) с высокоэнергетической связью во втором атоме углерода и отщепляется молекула воды

10. Реакция субстратного фосфорилирования
В ходе десятой реакции фосфорильная группа переносится на ADP. При этом синтезируется АТР и пируват (пировиноградная кислота). Эта реакция также необратима, поскольку высокоэкзергонична.

11. Реакция обратимого восстановления пировиноградной кислоты до молочной кислоты (в анаэробных условиях)
Если после гликолиза следует аэробное расщепление, пируват мигрирует в матрикс митохондрий, где, взаимодействуя с коэнзимом-А, участвует в образовании ацетил-СоА. В анаэробных условиях пируват при участии NADH восстанавливается до лактата (молочной кислоты), который при этом является конечным продуктом гликолиза. Затем в аэробных условиях лактат может обратно превратиться в пируват и окислиться в митохондриях.

_______________
Источник информации:
1. Биология для поступающих в вузы / Г.Л. Билич, В.А. Крыжановский. — 2008.
2. Биология в таблицах и схемах / Спб. — 2004.
3. Биохимия в схемах и таблицах / И. В. Семак — Минск — 2011.
Гликолиз и глюконеогенез. Аэробное окисление глюкозы.
» data-shape=»round» data-use-links data-color-scheme=»normal» data-direction=»horizontal» data-services=»messenger,vkontakte,facebook,odnoklassniki,telegram,twitter,viber,whatsapp,moimir,lj,blogger»>
ГЛИКОЛИЗ. ГЛЮКОНЕОГЕНЕЗ. АЭРОБНОЕ ОКИСЛЕНИЕ ГЛЮКОЗЫ.
ГЛИКОЛИЗ
Гликолиз — это сложный ферментативный процесс расщепления глюкозы до двух молекул пирувата (аэробный гликолиз) или двух молекул лактата (анаэробный гликолиз, протекающий без потребления кислорода).
Суммарное уравнение анаэробного гликолиза:
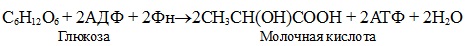
Гликолиз функционирует во всех живых клетках. Все ферменты локализованы в цитозоле, формируя полиферментный комплекс.
Гликолиз осуществляется в два этапа.
I. Подготовительный этап — дихотомический распад глюкозы на две молекулы глицеральдегид-3-фосфата. Превращения сопровождаются затратой 2 АТФ.
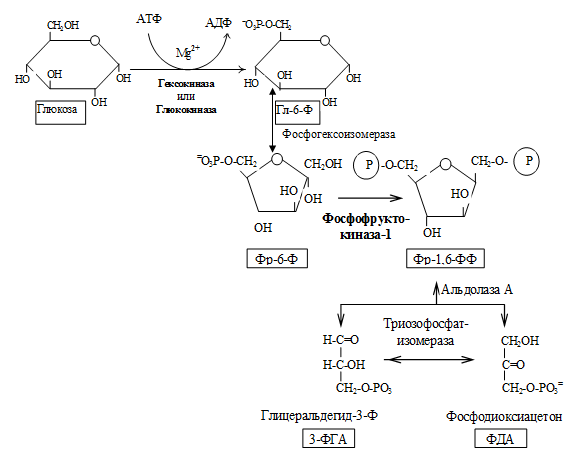
II. Этап гликолитической оксидоредукции — превращение глицеральдегид-3-фосфата в лактат. Включает окислительно-восстановительные реакции и реакции фосфорилирования, сопровождающиеся генерацией АТФ.
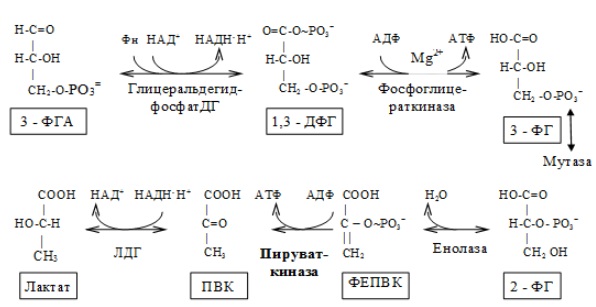
На втором этапе окисляются две молекулы глицеральдегид-3-фосфата, поэтому в реакциях впереди формулы субстрата следует ставить коэффициент 2.
В анаэробных условиях окисление НАДН . Н + , восстановленного в глицеральдегидфосфатдегидрогеназной реакции, происходит в лактатдегидрогеназной реакции. В аэробных условиях НАДН . Н + окисляется кислородом с участием ферментов дыхательной цепи, а выделяющаяся при этом энергия используется на синтез 1,5 или 2,5 моль АТФ (в зависимости от челночного механизма транспорта гликолитического НАДН . Н + в митохондрию).
Энергетический баланс гликолиза — две молекулы АТФ на одну молекулу глюкозы. На I этапе гликолиза расходуются две молекулы АТФ для активирования субстрата (в гексокиназной и фосфофруктокиназной реакциях). На II этапе образуются четыре молекулы АТФ (в фосфоглицераткиназной и пируваткиназной реакциях). Синтез АТФ осуществляется путем субстратного фосфорилирования.
Ключевые ферменты гликолиза:
1. Гексокиназа — это регуляторный фермент гликолиза во внепеченочных клетках. Гексокиназа аллостерически ингибируется глюкозо-6-фосфатом. Глюкокиназа — регуляторный фермент гликолиза в гепатоцитах. Синтез глюкокиназы индуцируется инсулином.
2. Фосфофруктокиназа-1. Это главный ключевой фермент, катализирует реакцию, лимитирующую скорость всего процесса (наиболее медленная реакция). Синтез фермента индуцируется инсулином. Аллостерические активаторы — АМФ, АДФ, фруктозо-2,6-дифосфат. Уровень фруктозо-2,6-дифосфата увеличивается под действием инсулина и понижается под действием глюкагона. Аллостерические ингибиторы — АТФ, цитрат.
3. Пируваткиназа. Фермент активен в нефосфорилированной форме. Глюкагон (в гепатоцитах) и адреналин (в миоцитах) стимулируют фосфорилирование фермента, а значит инактивируют фермент. Инсулин, наоборот, стимулирует дефосфорилирование фермента, а значит активирует фермент. Аллостерический активатор — Фр-1,6-ФФ. Аллостерический ингибитор — АТФ, ацетилКоА. Синтез фермента индуцирует инсулин.
Биологическая роль гликолиза:
1. Генерирование АТФ. Гликолиз — единственный процесс в клетках, продуцирующий АТФ без потребления кислорода. Клетки, имеющие мало или не имеющие вообще митохондрий, получают АТФ только в ходе гликолиза.
Значение гликолиза для эритроцитов. Гликолиз — единственный процесс, продуцирующий АТФ в эритроцитах и поддерживающий их целостность и функции.
Наследственный дефект пируваткиназы сопровождается гемолитической анемией. При этой патологии эритроциты имеют от 5 до 25 % нормальной пируваткиназной активности и, следовательно, скорость гликолиза низкая.
Промежуточный продукт гликолиза в эритроцитах — 2,3-дифосфоглицерат (2,3-ДФГ) — понижает сродство гемоглобина к кислороду, способствуя диссоциации кислорода из оксигемоглобина и переходу его в ткани. Нарушения гликолиза в эритроцитах могут оказывать влияние на транспорт кислорода. Так, при недостаточности гексокиназы наблюдается понижение уровня 2,3-ДФГ и ненормально высокое сродство гемоглобина к кислороду. И наоборот, при недостаточности пируваткиназы содержание 2,3-ДФГ вдвое превышает норму, что обусловливает низкое сродство гемоглобина к кислороду.
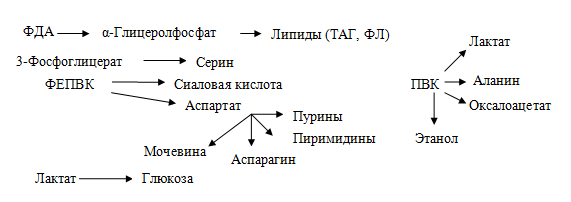
2. Является источником углеводородных радикалов для процессов биосинтеза в клетках:
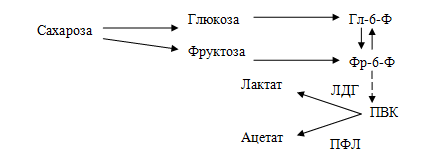
Патогенетическая взаимосвязь углеводов пищи и кариеса
Потребление легкоферментируемых углеводов, в частности сахарозы, инициирует кислотную деминерализацию эмали зубов. У бактерий имеются два альтернативных пути использования пирувата: первый — путь восстановления ПВК в молочную кислоту с участием лактатдегидрогеназы (ЛДГ), второй — расщепление ПВК на уксусную и муравьиную кислоту с участием пируватформиатлиазы (ПФЛ).
Схема расщепления сахарозы под влиянием ферментов бактерий полости рта:
Зависимость выхода Са 2+ из зубов от рН ротовой жидкости:
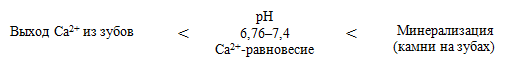
Аэробное окисление глюкозы
Это основной путь катаболизма глюкозы у аэробных организмов. Процесс осуществляется в три этапа. В аэробных условиях глюкоза окисляется до СО2 и Н2О.
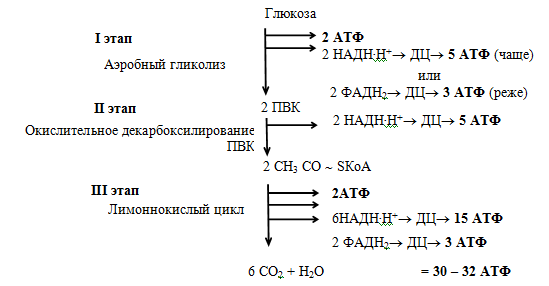
Энергетический баланс. Энергетический баланс аэробного окисления глюкозы —
30–32 моля АТФ на молекулу глюкозы.
ГЛЮКОНЕОГЕНЕЗ
Глюконеогенез — синтез глюкозы из соединений неуглеводной природы.
В организме взрослого человека за сутки может синтезироваться до 250 г глюкозы.
Глюконеогенез осуществляется главным образом в печени (синтезируетя до 90 % всей глюкозы), в корковом веществе почек и в энтероцитах (совсем незначительно).
Глюконеогенез стимулируется при длительном голодании, при ограничении поступления углеводов с пищей, в период восстановления после мышечной нагрузки, у новорождённых в первые часы после рождения.
Субстраты глюконеогенеза. Истинными субстратами глюконеогенеза являютя пируват, оксалоацетат, фосфодиоксиацетон, которые непосредственно включаются в этот процесс. Все вещества неуглеводной природы, дающие эти метаболиты, являются субстратами глюконеогенеза: лактат→ПВК, метаболиты цикла Кребса→ЩУК, глицерол→фосфодиоксиацетон, пропионил-КоА→метаболиты цикла Кребса→ЩУК, глюкогенные аминокислоты→ПВК или ЩУК. Главный источник субстратов глюконеогенеза — глюкогенные аминокислоты. К глюкогенным аминокислотам относятся все протеиногенные аминокислоты, кроме лейцина и лизина.
2ПВК + 4АТФ + 2ГТФ + 2НАДН . Н + + 2Н + 6Н2О → Глюкоза + 4АДФ + 2ГДФ + 6Фн + 2НАД +
Глюконеогенез протекает, в основном, по тому же пути, что и гликолиз, но в обратном направлении. Для обхода трех ключевых реакций гликолиза используются четыре специфических фермента глюконеогенеза.
Ключевые ферменты и ключевые реакции глюконеогенеза:
1. Пируваткарбоксилаза

2. Фосфоенолпируваткарбоксикиназа
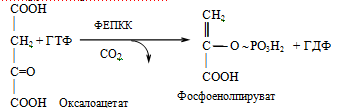
3. Фруктозо-1,6-бисфосфатаза
Фруктозо-1,6-бисфосфат + Н2О Фруктозо-6-фосфат + ФН
4. Глюкозо-6-фосфатаза
Глюкозо-6-фосфат + Н2О Глюкоза + ФН
Энергетический баланс. На синтез молекулы глюкозы из двух молекул пирувата расходуется 4АТФ и 2ГТФ (6АТФ). Энергию для глюконеогенеза поставляет процесс β-окис-ления жирных кислот.
Регуляция глюконеогенеза.
Глюконеогенез стимулируется в условиях гипогликемии при низком уровне инсулина и преобладании его антагонистов (глюкагона, катехоламинов, глюкокортикоидов).
1. Регуляция активности ключевых ферментов:
- фруктозо-1,6-бисфосфатаза по аллостерическому механизму активируется АТФ, ингибирутся Фр-1,6-ФФ и АМФ;
- пируваткарбоксилаза активируется СН3СО
КоА (аллостерический активатор).
2. Регуляция количества ключевых ферментов: глюкокортикоиды и глюкагон индуцируют синтез ключевых ферментов, а инсулин — репрессирует.
3. Регуляция количества субстрата: количество субстратов глюконеогенеза увеличивается под действием глюкокортикоидов (катаболическое действие на белки мышечной и лимфоидной ткани, на жировую ткань), а также глюкагона (катаболическое действие на жировую ткань).
Биологическая роль глюконеогенеза:
1. Поддержание уровня глюкозы в крови. При длительном голодании (голодание более суток) глюконеогенез является единственным процессом, поставляющим глюкозу в кровь.
2. Возвращение лактата в метаболический фонд углеводов. Лактат, образующийся в процессе анаэробного окисления глюкозы в эритроцитах и скелетных мышцах, транспортируется кровью в печень и превращается в гепатоцитах в глюкозу. Это так называемый межорганный цикл Кори.
3. Предотвращение лактатного ацидоза, то есть в ходе глюконеогенеза лактат крови превращается в глюкозу.
ГЛИКОЛИЗ
Гликолиз (от греч. glycys – сладкий и lysis – растворение, распад) – это последовательность ферментативных реакций, приводящих к превращению глюкозы в пируват с одновременным образованием АТФ.
При аэробных условиях пируват проникает в митохондрии, где полностью окисляется до СО2 и Н2О. Если содержание кислорода недостаточно, как это может иметь место в активно сокращающейся мышце, пируват превращается в лактат.
Итак, гликолиз – не только главный путь утилизации глюкозы в клетках, но и уникальный путь, поскольку он может использовать кислород, если
последний доступен (аэробные условия), но может протекать и в отсутствие кислорода (анаэробные условия).
Анаэробный гликолиз – сложный ферментативный процесс распада глюкозы, протекающий в тканях человека и животных без потребления кислорода. Конечным продуктом гликолиза является молочная кислота. В процессе гликолиза образуется АТФ. Суммарное уравнение гликолиза можно представить следующим образом:

В анаэробных условиях гликолиз – единственный процесс в животном организме, поставляющий энергию. Именно благодаря гликолизу организм человека и животных определенный период может осуществлять ряд физиологических функций в условиях недостаточности кислорода. В тех случаях, когда гликолиз протекает в присутствии кислорода, говорят об аэробном гликолизе .
Последовательность реакций анаэробного гликолиза, так же как и их промежуточные продукты, хорошо изучена. Процесс гликолиза катализируется одиннадцатью ферментами, большинство из которых выделено в гомогенном, клисталлическом или высокоочищенном виде и свойства которых достаточно известны. Заметим, что гликолиз протекает в гиало-плазме (цитозоле) клетки.
Первой ферментативной реакцией гликолиза является фосфорилирование, т.е. перенос остатка ортофосфата на глюкозу за счет АТФ. Реакция катализируется ферментом гексокиназой:
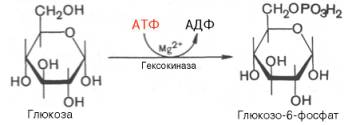
Образование глюкозо-6-фосфата в гексокиназной реакции сопровождается освобождением значительного количества свободной энергии системы и может считаться практически необратимым процессом.
Наиболее важным свойством гексокиназы является ее ингибирование глюкозо-6-фосфатом, т.е. последний служит одновременно и продуктом реакции, и аллостерическим ингибитором.
Фермент гексокиназа способен катализировать фосфорилирование не только D-глюкозы, но и других гексоз, в частности D-фруктозы, D-маннозы и т.д. В печени, кроме гексокиназы, существует фермент глюкокиназа, который катализирует фосфорилирование только D-глюкозы. В мышечной ткани этот фермент отсутствует (подробнее см. главу 16).
Второй реакцией гликолиза является превращение глюкозо-6-фос-фата под действием фермента глюкозо-6-фосфатизомеразы во фруктозо-6-фосфат:
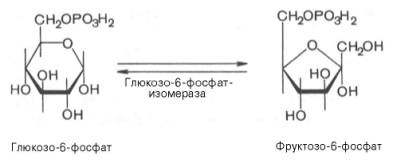
Эта реакция протекает легко в обоих направлениях, и для нее не требуется каких-либо кофакторов.
Третья реакция катализируется ферментом фосфофруктокиназой; образовавшийся фруктозо-6-фосфат вновь фосфорилируется за счет второй молекулы АТФ:
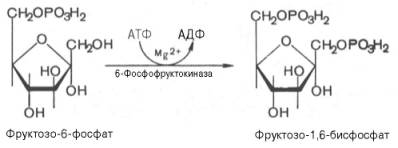
Данная реакция аналогично гексокиназной практически необратима, протекает в присутствии ионов магния и является наиболее медленно текущей реакцией гликолиза. Фактически эта реакция определяет скорость гликолиза в целом.
Фосфофруктокиназа относится к числу аллостерических ферментов. Она ингибируется АТФ и стимулируется АМФ . При значительных величинах отношения АТФ/АМФ активность фосфофруктокиназы угнетается и гликолиз замедляется. Напротив, при снижении этого коэффициента интенсивность гликолиза повышается. Так, в неработающей мышце активность фосфофруктокиназы низкая, а концентрация АТФ относительно высокая. Во время работы мышцы происходит интенсивное потребление АТФ и активность фосфофруктокиназы повышается, что приводит к усилению процесса гликолиза.
Четвертую реакцию гликолиза катализирует фермент альдолаза. Под влиянием этого фермента фруктозо-1,6-бисфосфат расщепляется на две фосфотриозы:
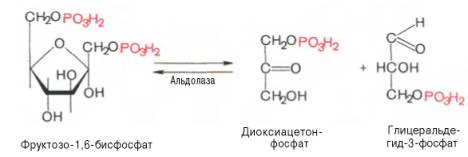
Эта реакция обратима. В зависимости от температуры равновесие устанавливается на различном уровне. При повышении температуры реакция сдвигается в сторону большего образования триозофосфатов (дигидро-ксиацетонфосфата и глицеральдегид-3-фосфата).
Пятая реакция – это реакция изомеризации триозофосфатов. Катализируется ферментом триозофосфатизомеразой:

Равновесие данной изомеразной реакции сдвинуто в сторону дигид-роксиацетонфосфата: 95% дигидроксиацетонфосфата и около 5% глице-ральдегид-3-фосфата. В последующие реакции гликолиза может непосредственно включаться только один из двух образующихся триозофосфатов, а именно глицеральдегид-3-фосфат. Вследствие этого по мере потребления в ходе дальнейших превращений альдегидной формы фосфотриозы ди-гидроксиацетонфосфат превращается в глицеральдегид-3-фосфат.
Образованием глицеральдегид-3-фосфата как бы завершается первая стадия гликолиза. Вторая стадия – наиболее сложная и важная. Она включает окислительно-восстановительную реакцию (реакция гликолитической оксидоредукции), сопряженную с субстратным фосфорилированием, в процессе которого образуется АТФ.
В результате шестой реакции глицеральдегид-3-фосфат в присутствии фермента глицеральдегидфосфатдегидрогеназы, кофермента НАД и неорганического фосфата подвергается своеобразному окислению с образованием 1,3-бисфосфоглицериновой кислоты и восстановленной формы НАД (НАДН). Эта реакция блокируется йод- или бромацетатом, протекает в несколько этапов:
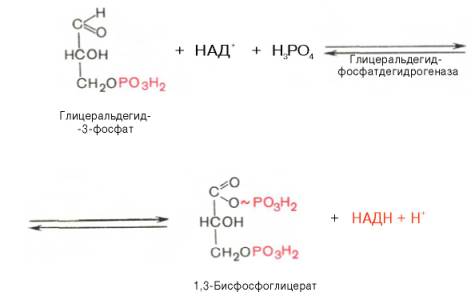
1,3-Бисфосфоглицерат представляет собой высокоэнергетическое соединение (макроэргическая связь условно обозначена знаком «тильда»
). Механизм действия глицеральдегидфосфатдегидрогеназы сводится к следующему: в присутствии неорганического фосфата НАД + выступает как акцептор водорода, отщепляющегося от глицеральдегид-3-фосфата. В процессе образования НАДН глицеральдегид-3-фосфат связывается с молекулой фермента за счет SH-групп последнего. Образовавшаяся связь богата энергией, но она непрочная и расщепляется под влиянием неорганического фосфата, при этом образуется 1,3-бисфосфоглицериновая кислота.
Седьмая реакция катализируется фосфоглицераткиназой, при этом происходит передача богатого энергией фосфатного остатка (фосфатной группы в положении 1) на АДФ с образованием АТФ и 3-фосфогли-цериновой кислоты (3-фосфоглицерат):
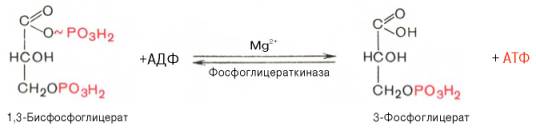
Таким образом, благодаря действию двух ферментов (глицеральде-гидфосфатдегидрогеназы и фосфоглицераткиназы) энергия, высвобождающаяся при окислении альдегидной группы глицеральдегид-3-фосфата до карбоксильной группы, запасается в форме энергии АТФ. В отличие от окислительного фосфорилирования образование АТФ из высокоэнергетических соединений называется субстратным фосфорилированием.
Восьмая реакция сопровождается внутримолекулярным переносом оставшейся фосфатной группы, и 3-фосфоглицериновая кислота превращается в 2-фосфоглицериновую кислоту (2-фосфоглицерат).
Реакция легкообратима, протекает в присутствии ионов Mg 2+ . Кофактором фермента является также 2,3-бисфосфоглицериновая кислота аналогично тому, как в фосфоглюкомутазной реакции роль кофактора выполняет глюкозо-1,6-бисфосфат:

Девятая реакция катализируется ферментом енолазой, при этом 2-фосфоглицериновая кислота в результате отщепления молекулы воды переходит в фосфоенолпировиноградную кислоту (фосфоенолпируват), а фосфатная связь в положении 2 становится высокоэргической:
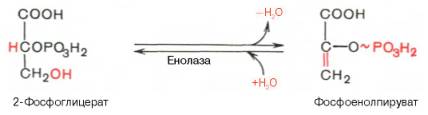
Енолаза активируется двухвалентными катионами Mg 2+ или Мn 2+ и ингибируется фторидом.
Десятая реакция характеризуется разрывом высокоэргической связи и переносом фосфатного остатка от фосфоенолпирувата на АДФ (субстратное фосфорилирование). Катализируется ферментом пируваткиназой:
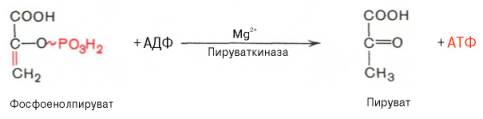
Для действия пируваткиназы необходимы ионы Mg 2+ , а также одновалентные катионы щелочных металлов (К + или др.). Внутри клетки реакция является практически необратимой.
В результате одиннадцатой реакции происходит восстановление пировиноградной кислоты и образуется молочная кислота. Реакция протекает при участии фермента лактатдегидрогеназы и кофермента НАДН, образовавшегося в шестой реакции:

Последовательность протекающих при гликолизе реакций представлена на рис. 10.3.
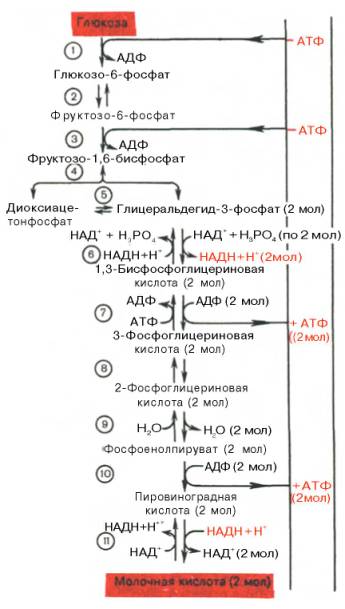
Рис. 10.3. Последовательность реакций гликолиза.
1 — гексокиназа; 2 — фосфоглюкоизоме-раза; 3 — фосфофруктокиназа; 4 — альдо-лаза; 5 — триозофосфатизомераза; 6 — гли-церальдегидфосфатдегидрогеназа; 7 -фосфоглицераткиназа; 8 — фосфоглицеромутаза; 9 — енолаза; 10 — пируватки-наза; 11 — лактатдегидрогеназа.
Реакция восстановления пирувата завершает внутренний окислительно-восстановительный цикл гликолиза. НАД + при этом играет роль промежуточного переносчика водорода от глицеральдегид-3-фосфата (6-я реакция) на пировиноградную кислоту (11-я реакция), при этом сам он регенерируется и вновь может участвовать в циклическом процессе, получившем название гликолитический оксидоредукции.
Биологическое значение процесса гликолиза заключается прежде всего в образовании богатых энергией фосфорных соединений. На первых стадиях гликолиза затрачиваются 2 молекулы АТФ (гексокиназная и фосфофрук-токиназная реакции). На последующих образуются 4 молекулы АТФ (фосфоглицераткиназная и пируваткиназная реакции). Таким образом, энергетическая эффективность гликолиза в анаэробных условиях составляет 2 молекулы АТФ на одну молекулу глюкозы.
Как отмечалось, основной реакцией, лимитирующей скорость гликолиза, является фосфофруктокиназная. Вторая реакция, лимитирующая скорость и регулирующая гликолиз – гексокиназная реакция. Кроме того, контроль гликолиза осуществляется также ЛДГ и ее изоферментами.
В тканях с аэробным метаболизмом (ткани сердца, почек и др.) преобладают изоферменты ЛДГ1 и ЛДГ2 (см. главу 4). Эти изоферменты инги-бируются даже небольшими концентрациями пирувата, что препятствует образованию молочной кислоты и способствует более полному окислению пирувата (точнее, ацетил-КоА) в цикле трикарбоновых кислот.
В тканях человека, в значительной степени использующих энергию гликолиза (например, скелетные мышцы), главными изоферментами являются ЛДГ5 и ЛДГ4. Активность ЛДГ5 максимальна при тех концентрациях пирувата, которые ингибируют ЛДГ1. Преобладание изоферментов ЛДГ4 и ЛДГ5 обусловливает интенсивный анаэробный гликолиз с быстрым превращением пирувата в молочную кислоту.
Как отмечалось, процесс анаэробного распада гликогена получил название гликогенолиза. Вовлечение D-глюкозных единиц гликогена в процесс гликолиза происходит при участии 2 ферментов – фосфорилазы а и фосфо-глюкомутазы. Образовавшийся в результате фосфоглюкомутазной реакции глюкозо-6-фосфат может включаться в процесс гликолиза. После образования глюкозо-6-фосфата дальнейшие пути гликолиза и гликогенолиза полностью совпадают:
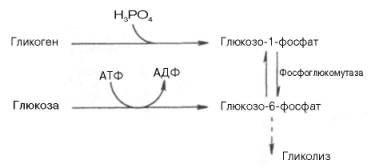
В процессе гликогенолиза в виде макроэргических соединений накапливаются не две, а три молекулы АТФ (АТФ не тратится на образование глюкозо-6-фосфата). Кажется, что энергетическая эффективность глико-генолиза выглядит несколько более высокой по сравнению с процессом гликолиза, но эта эффективность реализуется только при наличии активной фосфорилазы а. Следует иметь в виду, что в процессе активации фосфо-рилазы b расходуется АТФ (см. рис. 10.2).
http://farmf.ru/lekcii/glikoliz-i-glyukoneogenez-aerobnoe-okislenie-glyukozy/
http://xumuk.ru/biologhim/145.html