Дегидрогеназы, строение и действие
Окисление органических веществ в живых тканях растений и животных происходит с участием ферментов: дегидрогеназ – активаторов и переносчиков водорода (электрона) дыхательного материала и оксидаз – активаторов молекулярного кислорода.
Дегидрогеназы (от де. и новолат. hуdrogenium – водород), ферменты, катализирующие отщепление водорода от органических веществ. Коферментами Д. обычно являются динуклеотиды:
— никотинамидадениндинуклеотидфосфат (НАДФ) или флавинадениндинуклеотид (ФАД)
— флавинмононуклеотид (ФМН), которые акцептируют водород окисляемого вещества. Д. осуществляют первый этап биологического окисления.
Они играют большую роль в цикле Кребса, в гликолизе и в пентозофосфатном цикле. Некоторые Д., не связанные с коферментами, катализируют реакции окисления веществ непосредственно кислородом. Большая часть Д. в составе активного центра содержит металлы – цинк, марганец.
Дегидрогеназы действуют на протяжении всего процесса дыхания и по характеру действия делятся на аэробные и анаэробные. Первые переносят водород непосредственно на молекулярный кислород, а вторые – на какой-либо акцептор водорода. Дегидрогеназы относятся к двухкомпонентным ферментам – протеидам с пиридиновыми или флавиновыми (аллоксазиновыми) коферментами.
Пиридиновые дегидрогеназы относятся к группе анаэробных дегидрогеназ, которые являются акцепторами водорода субстрата. Специфичность их действия заключается в способности гидрирования и дегидрирования пиридиновых ядер.
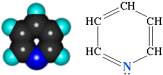
К флавопротеидным дегидрогеназам относятся ферменты, в состав которых фосфорный эфир рибофлавина.
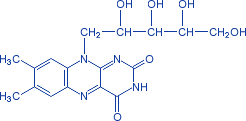
Рис. 2 − Рибофлавин
1. Березов Т. Т., Коровкин Б. Ф.Биологическая химия. М.: Медицина, 1990. С. 213–220; 1998. С. 305–317.
2. Николаев А. Я.Биологическая химия. М.: Высшая школа, 1989. С. 199–221.
3. Филиппович Ю. Б. Основы биохимии. М.: Высшая школа, 1993. С. 403–438.
4. Марри Р. и др. Биохимия человека. М.: Мир, 1993. Т. 1. С. 111–139.
5. Ленинджер А.Основы биохимии. М.: Мир, 1985. Т. 2. С. 403–438, 508–550.
6. АлбертсБ.идр.,Молекулярнаябиология клетки. М.: Мир, 1994.Т. 1. С. 430–459.
7. Скулачев В.П. Энергетика биологическихмембран. М.: Наука. 1989.
8. Это процесс потребление клетками тканей организма кислорода, который участвует в биологическом окислении. Такой вид окисления называют аэробным окислением. Если конечным акцептором в цепи переноса водорода выступает не кислород, а другие вещества (например пировиноградная кислота), то такой тип окисления называют анаэробным.
9. Т.о. биологическое окисление — это дегидрирование субстрата с помощью промежуточных переносчиков водорода и его конечного акцептора. Биологическое окисление питательных веществ происходит в митохондриях. В них были обнаружены ферменты, участвующие в цикле лимонной кислоты, дыхательной цепи, окислительного фосфорилирования, в расщеплении жирных кислот и ряда аминокислот.
10. Дыхательная цепь (ферменты тканевого дыхания) — это переносчики протонов и электронов от окисляемого субстрата на кислород. Окислитель — это соединение, способное принимать электроны. Такая способность количественно характеризуется окислительно-восстановительным потенциалом по отношению к стандартному водородному электроду, рН которого равен 7,0. Чем меньше потенциал соединения, тем сильнее его восстанавливающие свойства и наоборот.
11. Особенности тканевого дыхания
12. Процесс тканевого дыхания нельзя считать тождественным процессам биологического окисления (ферментативным процессам окисления различных субстратов, протекающим в животных, растительных и микробных клетках), поскольку значительная часть таких окислительных превращений в организме происходит в анаэробных условиях, т.е. без участия молекулярного кислорода, в отличие от дыхания тканей.
Большая часть энергии в аэробных клетках образуется благодаря дыханию тканей, и количество образующейся энергии зависит от его интенсивности. Интенсивность Д. т. определяется скоростью поглощения кислорода на единицу массы ткани; в норме она обусловлена потребностью ткани в энергии. Интенсивность его наиболее высока в сетчатке глаза, почках, печени; она значительна в слизистой оболочке кишечника, щитовидной железе, яичках, коре головного мозга, гипофизе, селезенке, костном мозге, легких, плаценте, вилочковой железе, поджелудочной железе, диафрагме, сердце, скелетной мышце, находящейся в состоянии покоя. В коже, роговице и хрусталике глаза интенсивность тканевого дыхания невелика. Гормоны щитовидной железы, жирные кислоты и другие биологически активные вещества способны активизировать тканевое дыхание.
Интенсивность такого дыхания определяют полярографически или манометрическим методом в аппарате Варбурга. В последнем случае для характеристики используют так называемый дыхательный коэффициент — отношение объема выделившегося углекислого газа к объему кислорода, поглощенного определенным количеством исследуемой ткани за определенный промежуток времени.
13. Т. о. любое соединение может отдавать электроны только соединению с более высоким окислительно-восстановительным потенциалом. В дыхательной цепи каждое последующее звено имеет более высокий потенциал, чем предыдущее.
14. Дыхательная цепь
15. Дыхательная цепь состоит из: НАД — зависимой дегидрогеназы; ФАД- зависимой дегидрогеназы; Убихинона (КоQ); Цитохрмов b, c, a+a3 .
16. НАД-зависимые дегидрогеназы. В качестве кофермента содержат НАД и НАДФ. Пиридиновое кольцо никотинамида способно присоединять электроны и протоны водорода.
17. ФАД и ФМН-зависимые дегидрогеназы содержат в качестве кофермента фосфорный эфир витамина В2 (ФАД).
18. Убихинон (КоQ) отнимает водород у флавопротеидов и превращается при этом в гидрохинон.
19. Цитохромы — белки хромопротеиды, способные присоединять электроны, благодаря наличию в своем составе в качестве простетических групп железопорфиринов. Они принимают электрон от вещества, являющегося немного боле сильным восстановителем, и передают его более сильному окислителю. Атом железа связан с атомом азота имидазольного кольца аминоксилоты гистидина с одной стороны от плоскости порфиринового цикла, а с другой стороны с атомом серы метионина. Поэтому потенциальная способность атома железа в цитохромах к связыванию кислорода подавлена.
20. В цитохроме с порфириновая плоскость ковалентно связана с белком через два остатка цистеина, а в цитохромах b и а, она ковалентно не связано с белком.
21. В цитохроме а+а3 (цитохромоксидазе) вместо протопорфирина содержатся порфирин А, который отличатся рядом структурных особенностей. Пятое координационное положение железа занято аминогруппой, принадлежащей остатку аминосахара, входящего в состав самого белка.
22. В отличии от гема гемолгобина атом железа в цитохромах может обратимо переходить из двух в трехвалентное состояние это обеспечивает транспорт электронов.
23. Потребности тканей в кислороде и его запасы
Потребности тканей в кислороде зависят от функционального состояния клеток, входящих в ее состав. Скорость потребления кислорода обычно выражается в мл кислорода на 1 г веса в минуту. В состоянии покоя кислород относительно интенсивно поглощается миокардом, серым веществом головного мозга (в частности, корой головного мозга), печенью и корковым веществом почек. В тоже время скелетные мышцы, селезенка и белое вещество головного мозга в покое потребляют мало кислорода.
При увеличении активности какого-либо органа потребность его в кислороде увеличивается. При физической нагрузке потребление кислорода миокардом может увеличиться в 3 — 4 раза, а работающими скелетными мышцами — более чем в 20 — 50 раз по сравнению с покоем. Потребление кислорода почками возрастает при увеличении интенсивности реабсорбции ионов натрия.
Количество кислорода, которое клетки могут использовать для окислительных процессов, зависит от конвекционного переноса кислорода кровью и диффузии кислорода из капилляров в ткани. Поскольку единственным запасом кислорода в большинстве тканей служит его физически растворенная фракция, снижение поступления кислорода приводит к кислородному голоданию и к замедлению окислительных процессов.
Единственной тканью, в которой имеются запасы кислорода, является мышечная ткань. Роль депо кислорода играет пигмент миоглобин, способный обратимо связывать кислород. Однако содержание миоглобина в мышцах человека невелико, так, среднее содержание миоглобина в сердце составляет 4 мг/г. Поскольку 1 г миоглобина может связать примерно до 1,34 мл кислорода, запасы кислорода в сердце составляют около 0,005 мл кислорода на 1г ткани. Этого количества в условиях полного прекращения доставки кислорода к миокарду может хватить для того, чтобы поддерживать окислительные процессы лишь в течение примерно 3 — 4 секунд.
Миоглобин играет роль кратковременного депо кислорода. В миокарде кислород, связанный с миоглобином, обеспечивает окислительные процессы в тех участках, кровоснабжение которых на короткий срок снижается или полностью прекращается во время систолы.
В начальном периоде интенсивной мышечной нагрузки увеличенные потребности скелетных мышц в кислороде частично удовлетворяются за счет кислорода, высвобождающегося миоглобином. В дальнейшем возрастает мышечный кровоток, и поступление кислорода к мышцам вновь становится адекватным. Восполнение запасов оксимиоглобина является составной частью кислородного долга, который должен быть покрыт каждым мышечным волокном после окончания работы.
Кислородное голодание тканей
При ряде патологических состояний страдает снабжение тканей кислородом. В этих случаях энергетические потребности клеток могут в течение короткого времени удовлетворяться за счет ограниченных запасов энергии в виде АТФ и креатинфосфата, а также за счет анаэробного гликолиза. Однако эти источники энергии недостаточны и могут использоваться недолго, так как в анаэробных условиях резко возрастает потребность клеток в глюкозе, поступление которой обычно не может удовлетворять эту потребность, и во-вторых, в процессе гликолиза образуется большое количество лактата, который медленно удаляется из ткани для последующей переработки (например, для расщепления в печени, почках или миокарде, или для синтеза гликогена). При значительном недостатке кислорода содержание лактата в крови постоянно нарастает, что приводит к нереспираторному ацидозу. Когда рН внутриклеточной среды падает ниже уровня оптимальной активности ферментных систем, наступают выраженные нарушения клеточного метаболизма.
Основные причины, приводящие к кислородному голоданию (тканевой гипоксии), это понижение напряжения кислорода в артериальной крови (артериальная гипоксия), уменьшение кислородной емкости крови (анемия) и уменьшение кровоснабжения того или иного органа (ишемия).
NADН-дегидрогеназа содержит несколько центров типа Fe2S2 и Fe4S4. Атомы железа в таких центрах могут принимать и отдавать электроны поочерёдно, переходя в ферро- (Fe 2 +) и ферри-(Fe 3 +) состояния. От железо-серных центров электроны переносятся на кофермент Q (убихинон) (рис.).
Дегидрогеназы – ферменты класса оксидоредуктаз (пиридинзависимые, флавинзависимые, аэробные и анаэробные типы, физиология, биохимия)

Дегидрогеназы представляют собой энзимы класса оксидоредуктаз, которые катализируют реакции отщепления водорода (то есть протонов и электронов) от субстрата, который является окислителем, и транспортируют его на другой субстрат, который восстанавливается.
В зависимости от химической природы акцептора, с которым взаимодействуют дегидрогеназы, их делят на несколько групп:
- Анаэробные дегидрогеназы, которые катализируют реакции, в которых акцептором водорода является соединение, которое отличается от кислорода.
- Аэробные дегидрогеназы, которые катализируют реакции где акцептором водорода может быть кислород (оксидазы) или другой акцептор. Аэробные дегидрогеназы относятся к флавопротеинам, продукт реакции — перекись водорода.
- Дегидрогеназы, которые обеспечивают транспортировку электронов от субстрата к акцептору электронов. К этой группе дегидрогеназ принадлежат цитохромы дыхательной цепи митохондрий.
- Дегидрогеназы, которые катализируют прямое введение в молекулу субстрата окисляется, 1 или 2 атомов кислорода. Такие дегидрогеназы получили название оксигеназы.
Функцию первичных акцепторов атомов водорода, отщепляющихся от соответствующих субстратов, выполняют дегидрогеназы 2 типов:
- пиридинзависимые дегидрогеназы — содержат коферменты никотинамид (НАД +) или никотинамидадениндинуклеотидфосфат (НАДФ +).
- флавинзависимые дегидрогеназы, простетической группой которых является флавинадениндинуклеотид (ФАД) или флавинмононуклеотид (ФМН).
Коферменты НАДФ+ (или НАД+) с апоферментом связаны непрочно и поэтому могут в клетке находиться как в связанном с апоферментом состоянии, так и быть отделенными от белковой части.
Пиридинзависимые дегидрогеназы относятся к анаэробному типу — водорастворимые ферменты, окисляющие полярные субстраты. Реакции, катализируемой пиридинзависимыми дегидрогеназами, в общем виде приведены в таких уравнениях:
SH2 + НАДФ+ → S + НАДФН + Н+
SH2 + НАД+ → S + НАДН + Н+
Рабочей структурой в молекуле НАД + или НАДФ + является пиридиновое кольцо никотинамид, который присоединяет в ходе ферментативной реакции один атом водорода и один электрон (гидрид-ион), а второй протон поступает в реакционную среду. Пиридинзависимые дегидрогеназы очень распространены в живых клетках. Они отщепляющие протоны и электроны от многих субстратов, восстанавливая НАД + или НАДФ + и передавая в дальнейшем восстановительные эквиваленты на другие акцепторы. НАД-зависимые дегидрогеназы главным образом катализируют окислительно-восстановительные реакции окислительных путей метаболизма — гликолиза, цикла Кребса, β-окисления жирных кислот, дыхательной цепи митохондрий и др. НАД является главным источником электронов для цепи переноса электронов. НАДФ используется главным образом в процессах восстановительного синтеза (в синтезе жирных кислот и стероидов).
Флавинзависимые дегидрогеназы — флавопротеины, простетическими группами в которых ФАД или ФМН — производные витамина В2, которые прочно (ковалентно) связаны с апоферментом. Данные дегидрогеназы — мембраносвязанные ферменты, окисляющие неполярные и малополярные субстраты. Рабочей частью молекулы ФАД или ФМН, которая участвует в окислительно-восстановительных реакциях, является изоалоксазиновое кольцо рибофлавина, который акцептирует два атома водорода (2Н+ + 2е-) от субстрата.
Общее уравнение реакции с участием флавинзависимых дегидрогеназ выглядит так:
SH2 + ФМН → S + ФМН-Н2
SH2 + ФАД+ → S + ФАДН2
В процессах биологического окисления данные ферменты играют роль как анаэробных, так и аэробных дегидрогеназ. К, анаэробным дегидрогеназам принадлежит НАДН-дегидрогеназа, ФМН-зависимый фермент, который передает электроны от НАДН на более электроположительны компоненты дыхательной цепи митохондрий. Другие дегидрогеназы (ФАД-зависимые) переносят электроны непосредственно от субстрата на дыхательную цепь (например, сукцинатдегидрогеназа, ацил-КоА-дегидрогеназа). Транспортировка электронов от флавопротеинов к цитохромоксидазам в дыхательной цепи обеспечивают цитохромы, которые, кроме цитохромоксидазы, по классификации как анаэробные дегидрогеназы. Цитохромы – представляют собой железосодержащие протеины митохондрий — гемпротеины, которые за счет обратного изменения валентности геминового железа выполняют функцию транспортировки электронов в аэробных клетках непосредственно в цепях биологического окисления: цитохром (Fe3 +) + е → цитохром (Fe2 +).
В состав дыхательной цепи митохондрий входят цитохромы b, с1, с, а и а3 (цитохромоксидаза). Кроме дыхательной цепи, цитохромы содержатся в эндоплазматическом ретикулуме (450 и b5). К аэробным флавинзависимым дегидрогеназам принадлежат оксидазы L-аминокислот, ксантиноксидаза и др.
Дегидрогеназы, катализируют включение одного или двух атомов кислорода в молекулу субстрата, получили название оксигеназы. В зависимости от количества атомов кислорода, которые взаимодействуют с субстратом, оксигеназы делят на 2 группы:
Диоксигеназы катализируют присоединяет в молекулу субстрата 2 атомов кислорода: S + O2 → SO2. Это, в частности, негемовые железосодержащие ферменты, катализирующие реакции синтеза гомогентизиновой кислоты и ее окисления на пути катаболизма тирозина. Железосодержащая липооксигеназа катализирует включение О2 в арахидоновую кислоту, первую реакцию в процессе синтеза лейкотриенов. Пролин- и лизиндиоксигеназы катализируют реакции гидроксилирования остатков лизина и пролина в проколлагене. Монооксигеназы катализируют присоединение к субстрату только 1 из атомов молекулы кислорода. При этом, второй атом кислорода восстанавливается до воды:
SH + О2 + НАДФН + Н+
К монооксигеназам принадлежат ферменты, которые участвуют в обмене веществ многих лекарственных субстанций путем их гидроксилирования. Эти ферменты имеют локализацию преимущественно в микросомальной фракции печени, надпочечников, половых желез и других тканей. Поскольку чаще всего субстрат в монооксигеназных реакциях гидроксилируется, данную группу энзимов также называют гидроксилазы.
Монооксигеназы катализируют реакции гидроксилирования холестерина (стероидов) и превращение их в биологически активные субстанции, в том числе — половые гормоны, гормоны надпочечников, активные метаболиты витамина D — кальцитриол, а также реакции детоксикации путем гидроксилирования ряда токсичных веществ, лекарственных средств и продуктов их превращения для организма. Монооксигеназная мембранная система эндоплазматического ретикулума гепатоцитов содержит НАДФН + Н+, флавопротеины с кофактором ФАД, протеин (адренотоксин), содержащий негемовое железо, и гемпротеин — цитохром Р450. В результате гидроксилирования неполярных гидрофобных субстанций повышается их гидрофильность, что способствует инактивации биологически активных веществ или обезвреживанию токсичных субстанций и экскреции их из организма. Некоторые лекарственные субстанции, такие, как фенобарбитал, имеют способность индуцировать синтез микросомальных ферментов и цитохрома Р450.
Существуют монооксигеназы, которые не содержат цитохрома Р450. К ним относятся ферменты печени, которые катализируют реакции гидроксилирования фенилаланина, тирозина, триптофана.
Полезно знать
© VetConsult+, 2015. Все права защищены. Использование любых материалов, размещённых на сайте, разрешается при условии ссылки на ресурс. При копировании либо частичном использовании материалов со страниц сайта обязательно размещать прямую открытую для поисковых систем гиперссылку, расположенную в подзаголовке или в первом абзаце статьи.
Биологическое окисление. цикл Кребса, этапы, дыхательная цепь
» data-shape=»round» data-use-links data-color-scheme=»normal» data-direction=»horizontal» data-services=»messenger,vkontakte,facebook,odnoklassniki,telegram,twitter,viber,whatsapp,moimir,lj,blogger»>
Биологическое окисление
Указанное выше уравнение для окислительно-восстановительной реакции представляет собой обобщенную форму, так как изображает процесс окисления субстратов как прямое дегидрирование, причем кислород выступает в роли непосредственного акцептора водорода. На самом деле кислород участвует в транспорте электронов иным образом. Существуют промежуточные переносчики при транспорте электронов от исходного донора электронов SH2 к терминальному акцептору – О2. Полный процесс представляет собой цепь последовательных окислительно-восстановительных реакций, в ходе которых происходит взаимодействие между переносчиками. Каждый промежуточный переносчик вначале выступает в роли акцептора электронов и протонов и из окисленного состояния переходит в восстановленную форму. Затем он передает электрон следующему переносчику и снова возвращается в окисленное состояние. На последней стадии переносчик передает электроны кислороду, который затем восстанавливается до воды. Совокупность последовательных окислительно-восстановительных реакций называется цепью переноса (транспорта) электронов, или дыхательной цепью:

Перенос электронов и протонов с участием промежуточных переносчиков. SH2 – исходный донор протонов и электронов; P – промежуточные переносчики; E1, E2, E3, E4 – ферменты окислительно-восстановительных реакций
Промежуточными переносчиками в дыхательной цепи у высших организмов являются коферменты: NAD + (никотинамид-адениндинуклеотид), FAD и FMN (флавинадениндинуклеотид и флавинмононуклеотид), кофермент Q (CoQ), семейство гемсодержащих белков – цитохромов (обозначаемых как цитохромы b, С1, С, А, А3) и белки, содержащие негеминовое железо. Все участники этой цепи разделены на четыре окислительно-восстановительные системы, связанные убихиноном (CoQ) и цитохромом С. Процесс начинается с переноса протонов и электронов от окисляемого субстрата на коферменты NAD+ или FAD. Это определяется тем, является ли дегидрогеназа, катализирующая первую стадию, NAD – зависимой или FAD – зависимой. Если процесс начинается с NAD + , то следующим переносчиком будет FMN.

Последовательность промежуточных переносчиков протонов и электронов в дыхательной цепи
Тип участвующей дегидрогеназы зависит от природы субстрата. Но каким бы ни был исходный субстрат, электроны и протоны от флавинов переносятся к коферменту Q, а дальше пути электронов и протонов расходятся. Электроны с помощью системы цитохромов достигают кислорода, который затем, присоединяя протоны, превращается в воду. Чтобы разобраться в системе транспорта электронов, необходимо познакомиться с отдельными ее участниками. NAD – зависимая дегидрогеназа катализирует реакции окисления непосредственно субстрата (первичная дегидрогеназа). NAD + является коферментом и выполняет роль акцептора водорода:

Символ 2Н + означает два электрона и два протона, обычно переносимые в виде гидрид иона. В этом случае вместо терминов «донор электронов» и «акцептор электронов» иногда используют термины «донор или акцептор водорода». FAD – зависимая дегидрогеназа также выполняет функцию первичной дегидрогеназы. Коферментом является FAD, который является акцептором водорода от субстрата. NADH – дегидрогеназа катализирует окисление NADH и восстановление убихинона (CoQ). Переносчиком водорода является кофермент – FMN (комплекс 1). В процессе реакции водород сначала присоединяется к FMN, соединенному с ферментом, а затем передается на убихинон. Флавиновые коферменты (FAD и FMN) прочно связаны с ферментом как простетические группы, поэтому ферменты, в состав которых они входят, называются флавопротеины. Флавинмононуклеотид (FMN), или рибофлавин фосфат, неразрывно связан с белковой частью фермента. Строго говоря, FMN не является нуклеотидом, так как флавиновая часть связана с рибитолом, а не с рибозой.
Убихинон (кофермент Q) – производное изопрена:

Название «убихинон» возникло из-за его повсеместной распространенности в природе. Кофермент Q действует как переносчик электронов на цитохромы.
Цитохромы – это гемопротеины – белки, содержащие в качестве прочно связанной простетической группы гем:

Простетическая группа гема в структуре цитохромов
Атом железа в геме может менять валентность, присоединяя или отдавая электроны.В дыхательной цепи цитохромы служат переносчиками электронов и располагаются соответственно величине окислительно-восстановительного потенциала следующим образом: B, С1, С, а, а3. Гемовые группы цитохромов связаны с белковой частью донорно-акцепторными связями между ионом железа и соответствующими аминокислотными остатками:

Связывание гема с белковой частью цитохрома С
В цитохромах С и С1 дополнительные ковалентные связи формируются между тиогруппами цистеина и боковыми винильными группами гема. QН2-дегидрогеназа (комплекс III) представляет собой комплекс цитохромов b и С1. Этот фермент катализирует окисление восстановленного кофермента Q и перенос электронов на цитохром С. Электроны последовательно переносятся атомами железа цитохромов b и С1, а затем поступают на цитохром С. Протоны после окисления QH2 освобождаются в раствор.
Цитохромоксидаза включает комплекс цитохромов а и а3 (комплекс IV). Цитохромоксидаза кроме гема содержит ионы меди, которые способны менять валентность и таким способом участвовать в переносе электронов.
Цитохромоксидаза переносит электроны с цитохрома С на кислород. В переносе электронов участвуют сначала ионы железа цитохромов а и а3, а затем ион меди цитохрома а3. Молекула кислорода связывается с железом в геме цитохрома а3. Следовательно, переход электронов на кислород с иона меди цитохрома а3, происходит на молекуле фермента. Каждый из атомов молекулы кислорода присоединяет по два электрона и протона, образуя при этом молекулу воды.
Белки, содержащие негеминовое железо. Некоторое количество атомов железа в митохондриях связано не в геме цитохромов, а образует комплексы с другими белками. Эти белки называют также железосерными, так как атомы железа связаны с атомами серы цистеиновых остатков. Белки, содержащие негеминовое железо, участвуют в переносе электронов на нескольких стадиях, однако, не совсем ясны их локализация и механизм действия.
Биологическое окисление. Синтез АТФ
Во внутренней мембране митохондрий расположен интегральный белковый комплекс – Н+-зависимая АТФ-синтаза seu Н+-зависимая АТФ-аза (два разных названия связаны с полной обратимостью катализируемой реакции), обладающий значительной молекулярной массой – более, чем 500кДа. Состоит из двух субъединиц: FO и F1.
F1 представляет из себя грибовидный вырост на матриксной поверхности внутренней митохондриальной мембраны, FO же пронизывает эту мембрану насквозь. В толще FO расположен протонный канал, позволяющий протонам возвращаться обратно в матрикс по градиенту их концентраций.
F1 способна связывать АДФ и фосфат на своей поверхности с образованием АТФ – без затраты энергии, но обязательно в комплексе с ферментом. Энергия необходима лишь для освобождения АТФ из этого комплекса. Эта энергия выделяется в результате тока протонов через протонный канал FO.
В дыхательной цепи сопряжение абсолютно: ни одно вещество не может окисляться без восстановления другого вещества.
Но при синтезе АТФ сопряжение одностороннее: окисление может идти без фосфорилирования, а фосфорилирование без окисления никогда не идёт. Это означает, что система МтО может работать без синтеза АТФ, но АТФ не может быть синтезирована, если не работает система МтО.
Биологическое окисление. Сопряжение работы дыхательной цепи с процессом синтеза АТФ
Существование такого сопряжения доказывается тем, что можно ингибировать образование АТФ, не нарушая процесса транспорта электронов. Это достигается добавлением химических веществ, названных разобщителями. После удаления разобщителей синтез АТФ восстанавливается.
Существует несколько гипотез, объясняющих механизм сопряжения. Одной из них является хемоосмотическая теория. Цепь транспорта электронов функционирует как протонная (Н + )помпа, осуществляя перенос протонов из матрикса через внутреннюю мембрану в межмембранное пространство. Эндоэргический процесс выброса протонов из матрикса возможен за счет экзоэргических окислительно-восстановительных реакций дыхательной цепи. Перенос протонов приводит к возникновению разности концентрации Н + с двух сторон митохондриальной мембраны: более высокая концентрация будет снаружи и более низкая – внутри. Митохондрия в результате переходит в «энергизованное» состояние, так как возникает градиент концентрации Н + и одновременно разность электрических потенциалов со знаком плюс на наружной поверхности.
Электрохимический потенциал способен совершать «полезную» работу, он заставляет протоны двигаться в обратном направлении, но мембрана непроницаема для них кроме отдельных участков, называемых протонными каналами. Обратный перенос протонов в матрикс является экзоэргическим процессом, высвобождающаяся при этом энергия используется на фосфорилирование ADP. Эту реакцию катализирует фермент Н + -АТФ-синтетаза, располагающаяся в области протонных каналов на внутренней поверхности внутренней мембраны.

С опряжение цепи транспорта электронов и фосфорилирования ADP посредством протонного градиента

С труктура компонентов комплекса I, обеспечивающего функционирование «протонной помпы» при окислении NADH
Биологическое окисление. ОСНОВНЫЕ ПРОЦЕССЫ, ДЛЯ КОТОРЫХ ИСПОЛЬЗУЕТСЯ ЭНЕРГИЯ АТФ:
1. Синтез различных веществ.
2. Активный транспорт (транспорт веществ через мембрану против градиента их концентраций). 30% от общего количества расходуемого АТФ приходится на Na+,К+-АТФазу.
3. Механическое движение (мышечная работа).
Биологическое окисление. Цикл Кребса
Цикл Кребса – цикл трикарбоновых кислот (ЦТК) является процессом окисления АцетилКоА – универсального продукта катаболизма углеводов, жиров и белков. Цикл Кребса протекает в митохондриях с участием 8 ферментов, которые локализованы в матриксе в свободном состоянии, или на внутренней поверхности внутренней мембраны. В Цикле Кребса участвуют 5 витаминов В1, В2, РР, пантотеновая кислота и липоевая кислота в виде коферментов тиаминпирофосфата, ФАД, НАД + , КоА и липоата.
Основной функции Цикла Кребса является образование водородных эквивалентов, которые в цепи окислительного фосфорилирования обеспечивают синтез макроэргических соединений.
Кроме того, ЦТК выполняет ведущую роль в процессах глюконеогенеза, переаминирования, дезаминирования АК, липогенеза и синтеза гема. Интегрирует все виды обмена веществ.
Регуляция Цикла Кребса. Осуществляется с участием 4 регуляторных ферментов: цитратсинтазы, изоцитрат ДГ, α-КГ ДГ и СДГ. Цикл Кребса ингибируется в основном НАДН2 и АТФ, которые являются продуктами ЦТК и цепи окислительного фосфорилирования. Активируют цикл Кребса в основном НАД + и АДФ.
Биологическое окисление. Реакции цикла Кребса
1). Цитратсинтаза локализуется в матриксе митохондрий, ее активируют ЩУК, НАД + ; ингибируют АТФ, НАДН2, Сукцинил-КоА, цитрат.

2). Аконитаза локализуется в матриксе митохондрий.

3). Окислительно-восстановительная реакция, самая медленная в ЦТК.
Изоцитратдегидрогеназа локализуется в матриксе митохондрий, ее активируют АМФ, Са 2+ , АДФ, НАД + ; ингибируют АТФ, НАДН2.

4). Окислительно-восстановительная реакция.
α-кетоглутаратдегидрогеназный комплекс состоит из 3 ферментов и содержит 5 коферментов: тиаминдифосфат, кофермент А, липоевая кислота, НАД + , ФАД.
α-КГ ДГ активируется Са 2+ , ингибируется сукцинил-КоА, АТФ, НАДН2.

5). Реакция субстратного фосфорилирования

6). Окислительно-восстановительная реакция.
Сукцинатдегидрогеназа, флавопротеин содержащий Fe2S2, связана с внутренней мембраной митохондрии. СДГ ингибирует ЩУК и Сукцинил-КоА,

7). Фумараза локализуется в матриксе митохондрий.

8). Окислительно-восстановительная реакция.
Малат ДГ локализуется в матриксе митохондрий.

Образовавшиеся молекулы ЩУК реагируют с новой молекулой Ацетил-КоА и цикл повторяется вновь.
Биологическое окисление. Энергетический баланс одного оборота ЦТК
В 4 окислительно-восстановительных реакциях ЦТК образуются 3 НАДН2 и 1 ФАДН2, которые направляются далее в дыхательную цепь окислительного фосфорилирования. В процессе окислительного фосфорилирования ДЦ из 1 НАДН2 образуется 3 АТФ, из 1 ФАДН2 – 2 АТФ. Из 1 ГТФ, образующейся в ЦТК за счет субстратного фосфорилирования, синтезируется 1 АТФ. Таким образом, за 1 цикл ЦТК из 3 НАДН2, 1 ФАДН2 и 1 ГТФ получается 12 АТФ.
Биологическое окисление. ЦИКЛ ТРИКАРБОНОВЫХ КИСЛОТ (ЦТК, лимоннокислый цикл, цикл Кребса)
ЦТК, как и реакции митохондриального окисления, протекает в митохондриях. Представляет собой серию реакций, замкнутых в цикл.


Образовавшиеся молекулы ЩУК реагируют с новой молекулой Ацетил-КоА и цикл повторяется вновь от образования цитрата до его превращения в ЩУК.
В реакциях этого цикла участвуют четыре из девяти субстратов МтО.
Происходит серия дегидрогеназных реакций. Из них 3-я, 4-я и 8-я происходят с участием НАД-зависимых дегидрогеназ, и каждая из этих реакций позволяет получить 3 молекулы АТФ. На 6-й стадии происходит ФАД-зависимая дегидрогеназная реакция, которая сопряжена с образованием 2-х молекул АТФ (Р/О = 2).
На 5-й стадии 1 молекула АТФ образуется путем субстратного фосфорилирования.
Итого за 1 оборот ЦТК образуется 12 молекул АТФ.
Смысл ЦТК заключается в том, чтобы остатки уксусной кислоты расщепились с образованием большого количества АТФ. Кроме того, из ацетатных остатков образуется СО2 и Н2О, как конечные продукты обмена веществ.
СО2 образуется в ходе ЦТК дважды:
1. на третьей стадии (окисление изоцитрата)
2. на четвертой стадии (окисление альфа-кетоглутарата).
Если прибавить еще 1 молекулу СО2, которая образуется до начала ЦТК – при превращении ПВК в Ацетил-КоА, то можно говорить о трех молекулах СО2, образующихся при распаде ПВК. Суммарно эти молекулы, образующиеся при распаде ПВК, составляют до 90% углекислоты, которая выводится из организма.
ИТОГОВОЕ УРАВНЕНИЕ ЦТК

БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ ЦТК
ГЛАВНАЯ РОЛЬ ЦТК – ОБРАЗОВАНИЕ БОЛЬШОГО КОЛИЧЕСТВА АТФ.
1. ЦТК – главный источник АТФ. Энергию для образования большого количества АТФ дает полный распад Ацетил-КоА до СО2 и Н2О.
2. ЦТК – это универсальный терминальный этап катаболизма веществ всех классов.
3. ЦТК играет важную роль в процессах анаболизма (промежуточные продукты ЦТК):
- – из цитрата → синтез жирных кислот
- – из aльфа-кетоглутарата и ЩУК → синтез аминокислот
- – из ЩУК → синтез углеводов
- – из сукцинил-КоА → синтез гема гемоглобина
АВТОНОМНАЯ САМОРЕГУЛЯЦИЯ ЦТК
В ЦТК два ключевых фермента:
- цитратсинтаза (1-я реакция)
- изоцитратдегидрогеназа (3-я реакция)
Оба фермента аллостерически ингибируются избытком АТФ и НАДН2. Изоцитратдегидрогеназа сильно активируется АДФ. Если АДФ нет, то этот фермент неактивен. В условиях энергетического покоя концентрация АТФ увеличивается, и скорость реакций ЦТК мала – синтез АТФ уменьшается.
Изоцитратдегидрогеназа ингибируется АТФ намного сильнее, чем цитратсинтаза, поэтому в условиях энергетического покоя повышается концентрация цитрата, и он выходит в цитоплазму по градиенту концентраций путем облегченной диффузии. В цитоплазме цитрат превращается в Ацетил-КоА, который участвует в синтезе жирных кислот.
Биологическое окисление. ЭНЕРГЕТИЧЕСКИЙ ЗАРЯД КЛЕТКИ (ЭЗК)
Многие процессы метаболизма регулируются соотношением АТФ/АДФ. Это соотношение характеризует энергетический статус (состояние) клетки (Энергетический Заряд Клетки).
Теоретически ЭЗК может меняться от 0 до 1. Обычно в норме величина ЭЗК поддерживается в клетке в пределах 0.85-0.90.
ОБРАТИТЕ ВНИМАНИЕ! Метаболические пути, приводящие к образованию АТФ, ингибируются высоким ЭЗК; метаболические пути, ведущие к использованию АТФ, активируются высоким ЭЗК.
Биологическое окисление. ЧЕЛНОЧНЫЕ МЕХАНИЗМЫ ПЕРЕНОСА ВОДОРОДА
Никотинамидные дегидрогеназы находятся не только в матриксе митохондрий, но и в цитозоле. Митохондриальная мембрана непроницаема для НАД, поэтому НАДН2, который образуется в цитозоле, может передать свой водород в митохондрию только с помощью специальных субстратных ЧЕЛНОЧНЫХ МЕХАНИЗМОВ. В митохондрию из цитозоля передается не сам НАДН2, а только водород, отнятый от него. Переносимый водород включается в молекулу вещества-челнока, способного проникать через митохондриальную мембрану. В митохондрии вещество-челнок отдает водород на митохондриальный НАД или ФАД и возвращается обратно в цитозоль.
 В клетках организма человека существуют 2 типа челночных механизмов.
В клетках организма человека существуют 2 типа челночных механизмов.
1. МАЛАТ-АСПАРТАТНЫЙ челнок (наиболее универсален для клеток организма). С высокой скоростью работает в миокарде, почечной ткани, печени.
В этой транспортной системе водород от цитоплазматического НАД передается на митохондриальный НАД(!), поэтому в митохондриях образуется 3 молекулы АТФ и не происходит потери энергии при переносе водорода. Для ткани печени малат-аспартатная система особенно важна, так как из митохондрии выводится Ацетил-КоА (в виде цитрата), а водород попадает в митохондрию (в составе малата).
 Таким образом, происходит не только челночный транспорт водорода от цитоплазматического НАД к митохондриальному, а и обратный транспорт Ацетил-КоА из митохондрий в цитоплазму в виде цитрата. В цитоплазме Ацетил-КоА может быть использован для синтеза жирных кислот.
Таким образом, происходит не только челночный транспорт водорода от цитоплазматического НАД к митохондриальному, а и обратный транспорт Ацетил-КоА из митохондрий в цитоплазму в виде цитрата. В цитоплазме Ацетил-КоА может быть использован для синтеза жирных кислот.
ЩУК может вернуться в цитоплазму и другим способом: она может вступить в реакцию трансаминирования с глутаминовой кислотой (СМОТРИТЕ РИСУНОК)
2. ГЛИЦЕРОФОСФАТНЫЙ челнок (встречается реже).
 В этой транспортной системе водород от цитоплазматического НАД передается на митохондриальный ФАД(!), и в митохондриях образуется 2 молекулы АТФ вместо 3-х – происходит потеря энергии при переносе водорода.
В этой транспортной системе водород от цитоплазматического НАД передается на митохондриальный ФАД(!), и в митохондриях образуется 2 молекулы АТФ вместо 3-х – происходит потеря энергии при переносе водорода.
В клетке существует не только челночный транспорт водорода от цитоплазматического НАД к митохондриальному. Происходит и обратный транспорт Ацетил-КоА из митохондрий в цитоплазму в виде цитрата. В цитоплазме Ацетил-КоА может быть использован для синтеза жирных кислот.
Биологическое окисление. Пути использования кислорода
Биологическое окисление. Историческая справка
Изучение окисления в организме было начато в 18 в. А. Лавуазье; в дальнейшем значительный вклад в исследование БО (его локализация в живых клетках, связь с др. процессами обмена веществ, механизмы ферментативных окислительно-восстановительных реакций, аккумуляция и превращение энергии и др.) внесли О. Варбург, Г. Виланд (Германия), Д. Кейлин, Х. Кребс, П. Митчелл (Великобритания), Д. Грин, А. Ленинджер, Б. Чанс, Э. Рэкер (США), а в СССР – А. Н. Бах, В. И. Палладин, В. А. Энгельгардт, С. Е. Северин, В. А. Белицер, В. П. Скулачев и др.
В 1777г французский химик Антуан Лоран Лавуазье (1743-1794), впервые правильно истолковал явление горения как процесс соединения веществ с О2. Так как горение и дыхание сопровождаются потреблением О2 и выделение СО2, он предположил что, в их основе лежит один процесс. Однако у дыхания были существенные отличия от горения: оно шло при низкой температуре, без пламени и в присутствии воды.
В конце XIX века русские исследователи А.Н. Бах и В.И. Палладин, работая независимо друг от друга, предложили 2 основные теории для объяснения процессов, протекающих в ходе биологического окисления.
В 1897г Алексей Николаевич Бах (1857-1946) сформулировал «перекисную теорию окисления», согласно которой молекула О2 сначала активируется. Активация О2 происходит в результате разрыва одной его связи (-О-О-) и присоединения к органическим веществам – оксидазам. Активированный О2 при взаимодействии с окисляемым веществом образует перекись.

Теория «перекисного окисления» Баха нашла свое подтверждение, однако главный механизм БО оказался иным.
Важнейшая заслуга в развитии учения о БО принадлежит русскому ученому ботанику и биохимику В.И. Палладину (1859-1922), который создал теорию «активации водорода». Он предположил, что окисление субстратов может происходить в 2 фазы:
1). Анаэробная фаза. В этой фазе особые вещества хромогены (R) отщепляют Н от субстратов и восстанавливаются (RH2).
2). Аэробная фаза. Восстановленные хромогены RH2 передают Н на О2.

В последствии теория В.И. Паладина подтвердилась для процессов митоходриального окисления, а хромогены, принимающие участие в отнятии водорода от субстратов, в настоящее время называются ферментами дегидрогеназами.
Биологическое окисление. Современные представления
Согласно современной теории БО:
- Окисление происходит как в аэробных, так и в анаэробных условиях.
- В аэробных организмах существует несколько путей использования О2.
- Реакции БО необходимы в первую очередь для получения энергии в форме АТФ, а также для синтеза новых веществ, разрушения ксенобиотиков и продуктов метаболизма.
- БО является сложным, многостадийным процессом, в котором ведущую роль играют ферменты оксидоредуктазы.
Окислительно-восстановительные реакции (ОВР) – реакции, в которых меняется степень окисления субстрата за счет присоединения/отщепления: 1) 1 е – ; 2) 2е – и 2Н + ; 3) атомов кислорода.
Биологическое окисление (БО) совокупность окислительно-восстановительных реакций, которые протекают во всех живых клетках.
Субстрат БО – вещество, способное отдавать электрон. (Любые вещества, способные вступать в реакции окисления).
Тканевое дыхание – окисление органических веществ в клетках, сопровождающееся потреблением О2 и выделением воды.
Субстрат тканевого дыхания – это вещество, которое отдает электрон непосредственно в цепь окислительного фосфорилирования. (Например, НАДН2, ФАДН2).
Дыхательная цепь – цепь переноса электронов. В переносе электронов от субстратов БО к О2 принимают участие: 1) НАД и НАДФ зависимые ДГ; 2) ФАД и ФМН зависимые ДГ; 3) цитохромы; 4) коэнзим Q; 5) белки, содержащие негеминовое железо.
Свободная энергия. Каждое органическое вещество обладает определенным запасом внутренней энергии (Е). Часть этой внутренней энергии может быть использована для совершения полезной работы, такую энергию называют свободной (G). Направление химической реакции определяется значением ∆G. У катаболических реакций ∆G отрицательно, эти реакции протекают самопроизвольно (экзергонические реакции). У анаболических реакций ∆G положительно, они протекают только при поступлении свободной энергии извне (эндергонические реакции).
Редокс-потенциалы. В каждой окислительно-восстановительной системе участвует окисленная и восстановленная формы одного соединения, которые образуют сопряженную окислительно-восстановительную или редокс-пару. Разные редокс-пары обладают различным сродством к электрону. Мерой сродства редокс-пары к электрону служит окислительно-восстановительный потенциал, или редокс-потенциал (Ео‘), величина которого прямо пропорциональна изменению свободной энергии ∆G. Величину Ео‘ выражают в вольтах; чем она отрицательнее, тем меньше сродство вещества к электронам и наоборот.
Самое низкое сродство к электрону -0,42В у водорода. Самое высокое сродство к электрону +0,82В у О2. Компоненты дыхательной цепи имеют редокс-потенциалы, занимающие промежуточное положение между -0,42В и +0,82В. В дыхательной цепи, вещества переносящие электрон, располагаются в порядке увеличения их редокс-потенциала. Электрон перемещается по дыхательной цепи от веществ с низким сродством к электрону к веществам с более высоким сродством к электрону, при этом происходит высвобождение свободной энергии, часть которой фиксируется в виде макроэргических соединений. Электроны в дыхательную цепь поставляют субстраты тканевого дыхания.
Биологическое окисление. Этапы унифицирования энергии пищевых веществ и образования субстратов тканевого дыхания
Образование субстратов тканевого дыхания осуществляется в несколько этапов:
1. При переваривании в ЖКТ происходит гидролиз полимеров (белков, полисахаридов) и ТГ до мономеров, которые потом всасывающихся в кровь и включающихся в промежуточный обмен.
2. В ходе катаболизма мономеры (моносахара, жирные кислоты и аминокислоты) превращаются в универсальное вещество – Ацетил-КоА (исключение некоторые АК).
3. Ацетил-КоА поступает в ЦТК, где из него в последовательных реакциях образуются субстраты тканевого дыхания: изоцитрат, α-КГ, сукцинат и малат.
4. Окисление субстратов тканевого дыхания сопровождается восстановлением коферментов НАДН2 и ФАДН2, которые затем отдают протоны в редокс-цепь окислительного фосфорилирования.
Биологическое окисление. Пути использования кислорода в клетке
В настоящее время выделено 4 основные пути использования кислорода в организме:
1. Оксидазный путь – окислительное фосфорилирование. Протекает в митохондриях, является основным источником АТФ в аэробных тканях. Потребляет 90% кислорода.
2. Монооксигеназный путь. Обеспечивает включение 1 атома кислорода в молекулу субстрата. Используется для синтеза новых веществ (стероидные гормоны), обезвреживания ксенобиотиков и токсических продуктов обмена в митохондриях и ЭПР.
3. Диоксигеназный путь. Обеспечивает включение молекулы кислорода в молекулу субстрата. Используется для деградации АК и синтеза новых веществ.
4. Пероксидазный и радикальный пути. Кислород участвует в образовании перекисей и активных радикалов, которые необходимы в пероксисомах для внутриклеточного пищеварения, разрушения макрофагами бактерий, вирусов, регуляции метаболизма и т.д. Перекиси и активные кислородные радикалы оказывают также повреждающее воздействие на структуры клеток и тканей, активируя ПОЛ. Разрушение перекисей и инактивация свободных радикалов осуществляется с помощью ферментативной и неферментативной антиокидантной системы.
Биологическое окисление. Типы оксидоредуктаз
В соответствие с путями потребления кислорода и катализируемыми реакциями, все оксидоредуктазы разделены на 5 групп:
1). Оксидазы удаляют Н из субстрата (S), используя в качестве акцептора Н только О2, продуктом реакции является вода. Все оксидазы содержат медь. Например, цитохромоксидаза, последний фермент дыхательной цепи.
2). Аэробные дегидрогеназы, или ФАД зависимые оксидазы, относятся к металлофлавопротеинам (ФАД, ФМН, Fe, Cu, Mo), находятся в пероксисомах и наружной мембране митоходрий. Они отнимают Н от субстрата, и передают его на О2 с образованием перекиси. Дегидрогеназа L-аминокислот (оксидаза L-аминокислот) в почках осуществляет окислительное дезаминирование. Ксантиндегидрогеназа (ксантиноксидаза) превращает пурины в мочевую кислоту. Моноаминоксидазы (МАО) окисляют гормон адреналин и некоторые биогенные амины. Диаминоксидазы (ДАО) окисляют гистамин и другие диамины и полиамины. Образующаяся перекись бактерицидное действие.
3). Анаэробные дегидрогеназы многочисленная группа, содержат в качестве коферментов НАДН2, НАДФН2, ФАД, ФМН, цитохромы. Они удаляют Н из субстрата, не используя в качестве акцептора протона О2. Анаэробные дегидрогеназы выполняют две главные функции: перенос Н с одного субстрата на другой в сопряженной ОВР и транспорт электронов в дыхательной цепи. Примеры ферментов: изоцитрат ДГ (НАДН2), сукцинат ДГ (ФАДН2), цитохромы в, с1, с, а и а3.
4). Оксигеназы. Включают кислород в молекулу субстрата. Работают в составе мультиферментного комплекса, содержащего ФАД зависимую ДГ, Fe2S2-белок, цитохромы Р450 или В5. Этот комплекс встроен в мембрану ЭПР или внутреннюю мембрану митохондрий.
а) Монооксигеназы. Обеспечивают включение 1 атома кислорода в молекулу субстрата. В качестве восстановителей (R) используется НАДФН2, НАДН2 и аскорбат. Участвуют в синтезе стероидных гормонов, обезвреживании ксенобиотиков. Фен-4-монооксигеназа окисляет фен в тир. Пролингидроксилаза (аскорбат) гидроксилирует пролин в проколлагене.
б) Диоксигеназы. Обеспечивают включение молекулы кислорода в молекулу субстрата, окисляют циклические соединения с разрывом цикла. Например, триптофаноксигеназа, превращает триптофан в фенилкенуренин.
5). Гидроксипероксидазы разрушают перекиси водорода и органические перекиси. Ферменты пероксидаза и каталаза. Пероксидазы, содержат протогем, восстанавливают перекись до воды за счет хинонов, аскорбата или цитохрома с. Глутатионперосидаза, содержит селен, восстанавливает перекись до воды за счет глутатиона.
Каталаза, гемопротеин, использует одну Н2О2 как донор е – , а вторую Н2О2 как акцептор е – , в результате образуется вода и молекула кислорода. Каталазы много в крови, костном мозге, слизистых оболочках, почках и печени.
Биологическое окисление. Варианты дыхательной цепи
1. ПОЛНАЯ ДЫХАТЕЛЬНАЯ ЦЕПЬ
В этой цепи окисляется небольшое количество субстратов, из которых главными являются четыре. Коэффициент Р/О=3.
| СУБСТРАТЫ | НИКОТИНАМИДНЫЕ ФЕРМЕНТЫ, ИХ ОКИСЛЯЮЩИЕ |
| изолимонная кислота (изоцитрат) | Изоцитратдегидрогеназа |
| Яблочная кислота (малат) | Малатдегидрогеназа |
| Глутаминовая кислота (глутамат) | Глутаматдегидрогеназа |
| бета-гидроксиацил-КоА | бета-гидроксиацил-КоА-дегидрогеназа |
Все ферменты полной цепи являются НАД-зависимыми дегидрогеназами.
2. СОКРАЩЕННАЯ (УКОРОЧЕННАЯ) ДЫХАТЕЛЬНАЯ ЦЕПЬ
Известны два субстрата. Главным из них является сукцинат (янтарная кислота).

Сукцинатдегидрогеназа – это комплекс II, который в укороченном варианте цепи является начальным звеном окисления. В составе комплекса – простетическая группа ФАД и FeSII. От ФАД.Н2 два атома водорода переносятся на KoQ. Значит, первое звено, которое имеется в полной цепи – исключается. Перепад окислительно-восстановительного потенциала между ФАД и KoQ невелик. Поэтому переноса H+ в межмембранное пространство в этой точке не происходит. DmH+ также создается, но меньший, чем в полной цепи. Значит, меньше и эффективность фосфорилирования – коэффициент Р/О=2.
Аналогичным образом окисляется и второй субстрат – ацил-КоА (активная форма любой жирной кислоты):

3. МАКСИМАЛЬНО СОКРАЩЕННАЯ (МАКСИМАЛЬНО УКОРОЧЕННАЯ) ДЫХАТЕЛЬНАЯ ЦЕПЬ.
Она представлена только цитохромной частью. Эксперименты показали, что здесь может быть окислен только один субстрат – аскорбиновая кислота, с участием фермента, восстановленные эквиваленты включаются в цепь на уровне цитохрома С цитохромоксидазы (цитохром аа3), но в реальных условиях такого окисления практически не происходит. Образуется вода и 1 молекула АТФ. Коэффициент Р/О=1.
Возможность такого окисления доказана в эксперименте in vitro. А в живой клетке аскорбиновая кислота обычно используется как донор водорода в системе окисления оксигеназного типа (реакции, катализируемые монооксигеназами: смотрите лекцию «Внемитохондриальное окисление»). Такие реакции с участием витамина «С» особенно важны для формирования белка коллагена, в котором за счет монооксигеназной реакции образуется гидроксипролин.
Биологическое окисление. ОКИСЛИТЕЛЬНОЕ ДЕКАРБОКСИЛИРОВАНИЕ ПИРОВИНОГРАДНОЙ И a-КЕТОГЛУТАРОВОЙ КИСЛОТ В МИТОХОНДРИЯХ
Этот вариант дыхательной цепи удлинен по сравнению с полной цепью за счет того, что первое звено катализируется не никотинамидным ферментом, а мультиферментным комплексом. Это единая надмолекулярная структура. В состав этого комплекса входят 3 фермента и 5 коферментов. Такой комплекс называется мультиферментным комплексом окислительного декарбоксилирования a-кетокислот, и он окисляет два субстрата:
1. Пировиноградную кислоту (пируват, ПВК)
Окисляется с помощью ферментов пируватдегидрогеназного комплекса;
2. a-кетоглутаровую кислоту (a-КГ)
Окисляется с помощью ферментов a-кетоглутаратдегидрогеназного комплекса.
Оба комплекса ферментов работают одинаково. Они катализируют реакции окислительного декарбоксилирования соответствующей a-кетокислоты.
Биологическое окисление. ОКИСЛИТЕЛЬНОЕ ДЕКАРБОКСИЛИРОВАНИЕ ПИРУВАТА
1-ю реакцию катализирует фермент ПИРУВАТДЕКАРБОКСИЛАЗА (Е1).
Простетической группой пируватдекарбоксилазы является тиаминдифосфат (ТПФ, тиаминпирофосфат, ТДФ) – это активная форма витамина В1. Активная часть ТПФ – тиазоловое кольцо и атом водорода в нем. Для краткости записывают: НС-ТПФ.

Пируватдекарбоксилаза отщепляет CO2, а оставшаяся оксиэтильная группа присоединяется к ТПФ.


2-ю и 3-ю реакцию процесса катализирует фермент АЦИЛТРАНСФЕРАЗА (Е2). Простетическая группа ацетилтрансферазы – амид липоевой кислоты. Катализирует перенос оксиэтильного остатка на свой собственный кофермент (строение ЛК знать описательно по учебнику). В ее составе есть дисульфидная связь.

На этом этапе уже произошло окисление оксиэтильного остатка до остатка уксусной кислоты, одновременно с этим началось восстановление липоевой кислоты.
3 этап – продолжает работать фермент ацилтрансфераза.
На этой стадии фермент переносит остаток уксусной кислоты на молекулу кофермента А (КоА). В КоА содержится остаток пантотеновой кислоты (витамин В3). Активной частью КоА является остаток тиоэтаноламина и его -SH группа.
Выяснено, что этот фермент переносит не только остатки уксусной кислоты, но и остатки других кислот (нециклических). Поэтому его еще называют коферментом ацилирования.

Второй фермент данного комплекса является не только ацетилтрансферазой, но и окислительным ферментом одновременно (дегидрогенизирующая ацетилтрансфераза).
Энергия этого окисления аккумулируется в виде макроэргической связи активной формы уксусной кислоты (ацетил-КоА).
4 этап катализируется ферментом ДИГИДРОЛИПОИЛДЕГИДРОГЕНАЗОЙ.
Кофермент этого этапа – особый ФАД с низким окислительно-восстановительным потенциалом (обозначается как ФАД’). Переносит протоны и электроны с дигидролипоата на НАД – последний кофермент в описанном комплексе.

В итоге можем записать суммарное уравнение:

Аналогично действует ферментный комплекс для окислительного декарбоксилирования a-кетоглутарата. Но названия двух из трех ферментов другие:
1-й фермент – декарбоксилаза a-кетоглутаровой кислоты
2-й фермент – сукцинилтрансфераза (переносит остаток янтарной кислоты)
Суммарное уравнение для этого комплекса:

В результате действия этих двух комплексов, каждый из двух приведенных субстратов (ПВК и a-КГ) – теряют СО2, два протона и два электрона, которые передаются на кислород с образованием Н2О и параллельно происходит фосфорилирование трех молекул АТФ. Сами субстраты превращаются в итоге в активную форму соответствующей кислоты.
Макроэргическая связь в молекулах этих активных форм может легко трансформироваться в макроэргическую связь молекулы АТФ. Например, для молекулы сукцинил-КоА:

Такой способ образования АТФ, когда нет сопряжения с процессами окисления, называется СУБСТРАТНЫМ ФОСФОРИЛИРОВАНИЕМ (при этом используется макроэргическая связь в молекуле какого-нибудь субстрата). В организмах высших животных и человека этот путь получения АТФ не является главным. Благодаря такому механизму окислительное декарбоксилирование a-кетокислот может считаться энергетически более выгодной, потому что образуется вещество с макроэргической связью – ацетил-КоА. Некоторые синтетазы используют макроэргические связи из Ацетил-КоА или Сукцинил-КоА для реакций биосинтеза.
Например, при синтезе гема гемоглобина используется часть молекул сукцинил-КоА:

Большая часть будет все же использоваться для синтеза ГТФ.
Другой продукт окислительного декарбоксилирования (Ацетил-КоА) может расходоваться в реакциях синтеза жирных кислот, холестерина, стероидов. А самая большая часть этого вещества используется при синтезе лимонной кислоты в реакциях Цикла Трикарбоновых Кислот.
Биологическое окисление. ЭНЕРГЕТИЧЕСКИЙ ИТОГ УДЛИНЕННОЙ ЦЕПИ.
Образуются 3 молекулы АТФ путем окислительного фосфорилирования. Продукты окислительного декарбоксилирования (ацетил-КоА и сукцинил-КоА) содержат макроэргическую связь. Энергия макроэргической связи сукцинил-КоА используется двумя способами.
- Субстратное фосфорилирование в ЦТК (так расходуется большая часть этого вещества).
- Реакции синтеза гема (меньшая часть сукцинил-КоА).
Энергия макроэргической связи Ацетил-КоА используется только в реакциях синтеза:
- Синтез лимонной кислоты (цитрата).
- Синтез кетоновых тел
- Синтез жирных кислот
- Синтез холестерина
Среди субстратов МтО почти нет веществ, которые организм получает с пищей – только глутаминовая кислота.
Субстраты МтО – в основном органические кислоты, которые образуются в организме в процессе катаболизма. В процессе катаболизма из большого количества разнообразных пищевых веществ образуется всего 2 вида общих метаболитов: Ацетил-КоА и ПВК.
Биологическое окисление. Пути использования кислорода
Биологическое окисление. ОКСИГЕНАЗНЫЙ ПУТЬ
Оксигеназы это ферменты, которые катализируют включение кислорода в молекулу субстрата. Оксигеназы работают в составе мультиферментных комплексов, встроенных в мембрану и участвуют в синтезе и деградации многих типов метаболитов. По способу включения кислорода их делят на: монооксигеназы и диоксигеназы.
Монооксигеназные реакции
Монооксигеназы это ферменты, которые включают в субстрат только один атом молекулы кислорода. Другой атом кислорода восстанавливается до воды с участием электронов и протонов НАДФН2, НАДН2, реже витамин С:
Монооксигеназные реакции протекают на цитоплазматической поверхности гладкого ЭПР, их называют микросомальным окислением, и на внутренней поверхности внутренней мембраны митохондрии.
1. Микросомальные монооксигеназные системы
Катализируют низкоспецифичные реакции. Эти монооксигеназы функционируют в комплексе с различными ЦПЭ:
А. Цепь НАДФН2-Р450 редуктаза – Цитохром Р450
Донорами протонов и электронов для этой цепи являются НАДФН2.
Субстратами являются гидрофобные вещества экзогенного (лекарства, ксенобиотики) и эндогенного (стероиды, жирные кислоты и т.д.) происхождения.
Регуляция активности осуществляется индукцией синтеза ферментов. Открыто более 250 веществ-индукторов (барбитураты, спирты, кетоны, стероиды, ароматические углеводороды).

.НАДФН2-Р450 редуктаза. Цитозольный домен содержит 2 кофермента ФАД и ФМН, гидрофобный домен фиксирует фермент в мембране. НАДФН2-Р450 редуктаза переносит электроны с НАДФН2 на цитохром Р450.
Цитохром Р450 – интегральный гемопротеин, содержит простетическую группу гем, имеет участки связывания для О2 и субстрата. Открыто 150 генов, кодирующих различные изоформы цитохрома Р450. Каждая из изоформ Р450 имеет много субстратов и отличается от других изоформ Р450 только белковой частью.
Цитохром Р450 передает 2 электрона на 1 атом молекулы кислорода, который превращается в О 2- , при взаимодействии с 2 протонами О 2- дает воду. Второй атом молекулы кислорода включается в субстрат RH, образуя ROH.
Б. Цепь НАДН2-цитохром b5 редуктаза – Цитохром b5 – стеароил-КоА-десатураза
Донорами протонов и электронов для этой цепи являются НАДН2.

НАДН2-цитохром b5 редуктаза – двухдоменный белок, цитозольный домен содержит ФАД, гидрофобный домен фиксирует фермент в мембране. НАДН2-b5 редуктаза переносит электроны с НАДН2 на цитохром b5.
Цитохром b5. Цитозольный домен содержит гем, гидрофобный домен фиксирует фермент в мембране. Цитохром b5 может передавать свои электроны на различные ферменты (цитохром Р450, Стеароил-КоА-десатуразу и т.д.), образуя различные ЦПЭ, при этом он участвует в десатурации и элонгации жирных кислот, в синтезе холестерина, плазминогенов и церамида.
Стеароил-КоА-десатураза – интегральный фермент, содержит негеминовое железо. Катализирует образование 1 двойной связи между 9 и 10 атомами углерода в жирных кислотах. Стеароил-КоА-десатураза переносит электроны с цитохрома b5 на 1 атом кислород, при участии протонов этот кислород образует воду. Второй атом кислорода включается стеариновую кислоту с образованием её оксиацила, который дегидрируется до олеиновой кислоты.
Биологическое окисление. Митохондриальные монооксигеназные системы
Митохондриальные монооксигеназные системы локализованы на внутренней поверхности внутренней мембране митохондрий и катализируют высокоспецифичные реакции.
Компонентами этих систем могут быть: НАДН2-зависимые ФАД-содержащее редуктазы, Fe2S2-белки (адренодоксин), цитохромы Р450, b5, элонгазы и т.д.

Митохондриальные монооксигеназные системы:
· в коре надпочечников, семенниках, яичниках и плаценте участвуют в синтезе стероидных гормонов из холестерина (гидроксилирование по С22 и С20 при отщеплении боковой цепи и по положениям 11β и 18);
· в почках участвуют в синтезе 1,25-диоксихолекальциферола (Витамин Д3) из 25-гидроксихолекальциферола;
· в печени участвуют в синтезе желчных кислот из холестерина;
· в соединительной ткани участвуют в гидроксилировании пролина в оксипролин в молекуле коллагена (пролингидроксилазы). Донором протонов и электронов является витамин С. С оксипролином зрелый коллаген приобретает механическую прочность.
Биологическое окисление. Диоксигеназные реакции
Диоксигеназы это ферменты, которые включают в субстрат оба атома молекулы кислорода:
Таким путем окисляются циклические трудноокисляемые структуры, реакции идут с разрывом цикла. Диоксигеназные реакции протекают на цитоплазматической поверхности гладкого ЭПР.
Гомогентизатдиоксигеназа печени, содержит Fe 2+ , участвует в катаболизме тирозина:

L-триптофандиоксигеназа печени, содержит гем, участвует в катаболизме триптофана:

Биологическое окисление. ПЕРОКСИДАЗНЫЙ И РАДИКАЛЬНЫЙ ПУТИ ИСПОЛЬЗОВАНИЯ КИСЛОРОДА
Кислород – потенциально опасное вещество. Молекулярный О2 является бирадикалом (˙О::О˙), но он стабилен так как, 2 неспаренных е – внешней орбиты имеют параллельные спины, (подавляют радикальную активность друг друга).
Кислород в составе молекулы Н2О стабилен, потому что его внешняя электронная орбита укомплектована е – (Н:О:Н). Химические соединения, в составе которых кислород имеет промежуточную степень окисления, имеют высокую реакционную способность и называются активными формами кислорода (АФК). К активным формам кислорода относятся свободные радикалы кислорода и перекиси. Свободный радикал – свободный атом или частица с неспаренным электроном.
Образование активных форм кислорода
АФК во многих клетках образуются в основном в ферментативных и неферментативных реакциях в результате последовательного присоединения е – к кислороду:
1) О2 + 1е – → О ∙ 2 супероксидный анион-радикал (˙О::О:).
2) О ∙ 2 +1е – → О 2- 2 пероксидный анион (:О::О:), он быстро протонируется с образованием перекиси водорода О 2- 2 + 2Н + → Н2О2 (Н:О::О:Н)
3) Н2О2 + 1е – → НО ∙ + ОН – гидроксильный радикал, ОН – протонируется с образованием воды ОН – + Н + → Н2О
4) ОН ∙ + 1е – → Н2О (Н:О:Н)
В организме донорами электронов являются:
1). ЦПЭ. Утечка электронов из ЦПЭ на кислород является основным путем образования активных форм кислорода в большинстве клеток:
a) В цепи окислительного фосфорилирования Q принимая 1 электрон превращается в свободный радикал семихинон НQ ∙ , который при реоксигенации ишемических тканей может непосредственно взаимодействовать с кислородом, образуя супероксидный анион-радикал: HQ· + O2 → Q+ О ∙ 2 + H + ;
b) в монооксигеназных реакциях электрон с цитохрома Р450 переходит на кислород с образованием супероксидного анион-радикала, который иногда теряется с активного центра.
c) Аэробные дегидрогназы (ФАД-зависимые оксидазы) переносят электроны и протоны с субстрата на кислород с образованием перекиси водорода. Примеры таких оксидаз — оксидазы аминокислот, супероксид дисмутаза, оксидазы, локализованные в пероксисомах.
2). Металлы переменной валентности. Наличие в клетках Fe 2+ или ионов других переходных металлов катализирует образования гидроксильных радикалов и других активных форм кислорода. Например, в эритроцитах окисление иона железа гемоглобина способствует образованию супероксидного анион-радикала.
Hb(Fe 2+ ) + O2 → MetHb(Fe 3+ ) + О ∙ 2
H2O2 + Fe 2+ → Fe 3+ + HO – + HO· (реакция Фентона)
HOCl + Fe 2+ → Fe 3+ + Cl – + HO· (реакция Осипова)
3). Радикалы. Активные формы кислорода, обмениваясь электроном, легко переходят друг в друга: О ∙ 2 + Н2О2 → О2 + НО ∙ + ОН –
Активные формы кислорода в организме также могут образовываться при гомолитическом разрыве связей под действием ионизирующего излучения. Ионизирующее излучение вызывает например, радиолиз воды с образованием Н2; Н2О2 и свободных радикалов: Н · , НО ∙ , О · . Это процесс в основном происходит на поверхности тела – в коже (понятие фотостарения).
Биологическое окисление. Ферментативные реакции образования АФК
Электроны, необходимые для образования АФК могут давать ЦПЭ. Утечка е – из ЦПЭ на кислород является основным путем образования АФК в большинстве клеток:
- В цепи окислительного фосфорилирования Q принимая 1 е – превращается в свободный радикал семихинон НQ ∙ , который при реоксигенации ишемических тканей может непосредственно взаимодействовать с кислородом, образуя супероксидный анион-радикал: HQ· + O2 → Q+ О∙2 + H + ;
- в монооксигеназных реакциях е – с цитохрома Р450 переходит на кислород с образованием супероксидного анион-радикала, который иногда теряется с активного центра.
- Аэробные дегидрогеназы (ФАД-зависимые оксидазы) переносят е – и Н + с субстрата на кислород с образованием перекиси водорода. Примеры таких оксидаз — оксидазы аминокислот, супероксид дисмутаза, оксидазы, локализованные в пероксисомах.
Неферментативные реакции образования АФК
Электроны, необходимые для образования АФК могут давать:
1). Металлы переменной валентности. Наличие в клетках Fe 2+ или ионов других переходных металлов катализирует образования АФК. Например, в эритроцитах окисление иона железа гемоглобина способствует образованию супероксидного анион-радикала.
Hb(Fe 2+ ) + O2 → MetHb(Fe 3+ ) + О ∙ 2
H2O2 + Fe 2+ → Fe 3+ + HO – + HO· (реакция Фентона)
HOCl + Fe 2+ → Fe 3+ + Cl – + HO· (реакция Осипова)
2). Радикалы. АФК, обмениваясь электроном, легко переходят друг в друга: О ∙ 2 + Н2О2 → О2 + НО ∙ + ОН –
АФК также могут образовываться в организме неферметативно при гомолитическом разрыве связей под действием ионизирующего излучения. Ионизирующее излучение вызывает например, радиолиз воды с образованием Н2; Н2О2 и свободных радикалов: Н · , НО ∙ , О · . Это процесс в основном происходит на поверхности тела – в коже (понятие фотостарения).
Биологическое окисление. Свойства активных форм кислорода
Кислородные радикалы, обладая высокой активностью, разрушают органические молекулы в реакциях свободно-радикального окисления (СРО). Большая часть этих реакций протекает с полиненасыщенными жирными кислотами липидов, и называется перекисным окислением липидов (ПОЛ). Реакции ПОЛ являются цепными.
Наиболее химически активным соединением является гидроксильный радикал – сильнейший окислитель. Время его жизни очень короткое (1 миллиардная доля секунды), но за это время он мгновенно вступает в цепные окислительные реакции в месте своего образования.
Супероксидный анион-радикал и перекись водорода более стабильные вещества, могут диффундировать от места образования, проникать через мембраны клеток. Однако, перекись водорода способствует образованию гидроксильного радикала по следующей реакции:
Fe 2+ + Н2О2 → Fe 3+ + НО ∙ + ОН –
Биологическое окисление. Использование активных форм кислорода в организме
1. Иммунная система. АФК используются фагоцитарными клетками – тканевыми макрофагами, моноцитами и гранулоцитами крови для разрушения бактерий, вирусов и онкоклеток.
- Фагоциты с участием НАДФН2-оксидазы выделяют супероксидный анион-радикал: НАДФН2 + 2O2 → НАДФ + + 2О∙2 + 2Н +
- Под действием супероксиддисмутазы (СОД) супероксидный радикал превращается в перекись водорода: 2О∙2 + 2H + → H2O2+ O2
- Под действием миелопероксидазы H2O2, превращается в гипохлорит – соединение, разрушающее стенки бактериальных клеток: H2O2 + Cl – → H2O + ClO – .
В условиях патологии могут произойти нарушения либо системы защитных ферментов (в частности, снижение активности СОД), либо ферментных систем, связывающих ионы железа в плазме крови (церулоплазмин и трансферрин) и в клетках (ферритин). В этом случае супероксидные радикалы и перекись водорода вступают в альтернативные реакции: Образование двухвалентного железа из трехвалентного: Fe 3+ + O •- → Fe 2+ + O2
Реакция перекиси водорода и гипохлорита с ионами двухвалентного железа:
При дефиците в клетках СОД, ферритина, а в плазме церулоплазмина, трансферрина активируются альтернативные реакции:
- Fe 3+ + О∙2 → Fe 2+ + O2
- Fe 2+ + H2O2 → Fe 3+ + НО ∙ + ОН –
- Fe 2+ + ClO – + H + → Fe 3+ + Cl – + НО ∙
Фактор некроза опухоли
2. Поддержание гомеостаза.
Эйказаноиды – медиаторы воспаления
3. Внутриклеточное пищеварение. В пероксисомах образуются АФК. Когда пероксисомы сливаются с фагосомами, АФК обеспечивают внутриклеточное пищеварение.
Биологическое окисление. Повреждающее действие активных форм кислорода в организме
Радикалы гидроксила химически исключительно активны и вызывают повреждение белков, нуклеиновых кислот и липидов биологических мембран. В белках происходит неферментативное окисление аминокислотных остатков гистидина, цистеина, триптофана. Таким образом, инактивируют многие ферменты. Особенно тяжелые последствия имеют повреждение ДНК и липидов. Радикалы НО ∙ вызывают разрыв нитей ДНК, обладая, в зависимости от ситуации, мутагенным, канцерогенным или цитостатическим действием. Радикалы гидроксила инициируют цепную реакцию ПОЛ, при этом нарушаются физико-химические свойства мембран – проницаемость, рецепторная функция и работа мембранных белков. Повреждение барьера приводит к нарушению регуляции внутриклеточных процессов и тяжелым расстройствам клеточных функций. Свободно-радикальные реакции часто вызывают гибель клеток и целом ускоряют процесс старения организма.
Биологическое окисление. Антиоксидантные клетки
Антиоксидантная система
В нормальных условиях процесс СРО находится под строгим контролем ферментативных и неферментативных систем клетки, от чего скорость его невелика. Химические соединения и физические воздействия, влияющие на скорость СРО, делят на прооксиданты и антиоксиданты.
Прооксиданты усиливают процессы СРО. Это высокие концентрации кислорода (например, при длительной гипербарической оксигенации больного), ферментные системы, генерирующие супероксидные радикалы (например, ксантиноксидаза, ферменты плазматической мембраны фагоцитов и др.), ионы двухвалентного железа.
Антиоксиданты тормозят СРО. Антиоксиданты, находящиеся в организме, образуют его ферментативную и неферментативную антиоксидантную систему.
1. Ферментативная антиоксидантная система
К ферментам, защищающим клетки от действия активных форм кислорода, относят супе-роксиддисмутазу, каталазу и глутатионпероксидазу. Наиболее активны эти ферменты в печени, надпочечниках и почках, где содержание митохондрий, цитохрома Р450 и пероксисом особенно велико.
Супероксиддисмутаза (СОД) превращает супероксидные анионы в перекись водорода:
Изоферменты СОД находятся и в цитозоле (Cu 2+ и Zn 2+ ) и в митохондриях (Mn 2+ ) и являются как бы первой линией защиты, потому что супероксидный анион образуется обычно первым из активных форм кислорода. СОД — индуцируемый фермент, т.е. синтез его увеличивается, если в клетках активируется СРО.
Каталаза – геминовый фермент, катализирует реакцию разрушения перекиси водорода. При этом образуется вода и молекулярный кислород:
Каталаза находится в основном в пероксисомах, где образуется наибольшее количество перекиси водорода, а также в лейкоцитах, где она защищает клетки от последствий «респираторного взрыва» и в эритроцитах, где она защищает гем гемоглобина от окисления.
Глутатионпероксидаза — обеспечивает разрушение перекиси водорода и гидропероксидов липидов при окислении глутатиона (у-глутамилцистеинилглицин): Н2О2 + 2 GSH → 2 Н2О + G-S-S-G. Глутатионпероксидаза в качестве кофермента содержит селен.
Глутатионредуктаза восстанавливает окисленный глутатион с участием НАДФН2:
GS-SG + НАДФН2 → 2 GSH + НАДФ + .
Недостаток глутатиона в клетках, например эритроцитах, который может быть обусловлен действием токсических веществ, например ионами тяжелых металлов или наследственным недостатком глутатионредуктазы приводит к активации перекисного окисления; это, в частности, наблюдается при некоторых видах гемолитических анемий.
Фосфолипаза в мембране отщепляет от фосфолипидов окисленные жирные кислоты, содержащие гидроперекисную группу (LOOH), тем самым разрушаются гидроперекиси липидов, предотвращается разветвление цепей окисления липидов в мембранах.
2. Неферментативная антиоксидантная система
“Липидные антиоксиданты” – производные фенола, способны инактивировать свободные радикалы в гидрофобном слое мембран и предотвращать развитие ПОЛ. К ним относится α-токоферол (витамин Е), убихинон (коэнзим Q), тироксин и синтетические соединения, например ионол (бутилированный гидрокситолуол).
Витамин Е (α-токоферол) самый распространённый липофильный антиоксидант, он обеспечивает защиту мембран от СРО. Различают 8 типов токоферолов, но α-токоферол наиболее активен. α-Токоферол отдаёт атом водорода радикалу липида ROO ∙ , восстанавливает его до гидропероксида (ROOH), а сам превращается в малоактивный свободный радикал, что прерывает ПОЛ:

Регенерацию α-токоферола осуществляет витамин С.
Свободный радикал витамина Е стабилен и не поддерживает ПОЛ, он взаимодействует с радикалами липидных перекисей, восстанавливает их, а сам превращается в стабильную окисленную форму — токоферолхинон.

Витамин С ингибирует СРО с помощью двух различных механизмов:
1). восстанавливает в мембранах токоферолхинон до витамина Е:
НО-аскорбат-ОН + α-ТФ-О· → α-ТФ-ОН + НО-аскорбат-О· (семидегидроаскорбиновая к-та)
НО-аскорбат-О· + α-ТФ-О· → α-ТФ-ОН + О=аскорбат=О (дегидроаскорбиновая к-та)
Регенерация аскорбиновой кислоты идет с участием ферментативных систем:
а). В микросомах, с участием комплекса НАДН2-редуктаза-цитохром b5:
2НО-аскорбат-О· + НАДН2 → 2НО-аскорбат-ОН + НАД +
б). В митохондриях, с участием НАДН2-семидегидроаскорбатредуктазы:
2НО-аскорбат-О· + НАДН2 → 2НО-аскорбат-ОН + НАД +
в). В цитозоле, с участием НАДФН2-дегидроаскорбатредуктазы:
О=аскорбат=О + 2НАДФН2 → НО-аскорбат-ОН + 2НАДФ +
г). В цитозоле, с участием GSH-дегидроаскорбатредуктазы:
О=аскорбат=О + 2GSH → НО-аскорбат-ОН + GS-SG
2). взаимодействует с активными формами кислорода — О ∙ 2, Н2О2, НО ∙ и инактивирует их.
β-Каротин, предшественник витамина А, также обладает антиоксидантным действием и ин-гибирует ПОЛ.
Соединения, связывающие железо. Большинство из них, включая такие природные соединения как дипептид карнозин, не просто связывают железо, но, самое главное, не дают ему возможности приникнуть в липидную фазу мембран, поскольку образующиеся комплексы, в силу своей полярности, не проникают в гидрофобную зону.
Для детоксикации двухвалентного железа в организме существует, по-видимому, целая система окисления и связывания ионов железа. В плазме крови эта система представлена ферментом церрулоплазмином (феррооксидазой), который окисляет Fe 2+ до Fe 3+ кислородом без образования свободных радикалов, и белком трансферрином, который связывает и переносит в кровяном русле ионы трехвалентного железа, которые затем захватывается клетками. В клетках железо может восстанавливаться аскорбиновой кислотой и другими восстановителями, но затем окисляется и депонируется в окисленной форме внутри ферментного белкового комплекса ферритина.
http://vetconsultplus.ru/%D0%94/Fermenty-degidrogenazy-kofermenty-NAD-NADF-FAD.html
http://farmf.ru/lekcii/biologicheskoe-okislenie-cikl-krebsa-chelnochnye-mexanizmy-perenosa-vodoroda/