Энергетический обмен
Обмен веществ
Обмен веществ (метаболизм) складывается из процессов расщепления и синтеза — диссимиляции и ассимиляции, постоянно протекающих в организме. Чтобы жизнь продолжалась, количество поступающей энергии должно превышать (или как минимум равняться) количеству расходуемой энергии, поэтому диссимиляция и ассимиляция поддерживают определенный баланс друг с другом.

Энергетический обмен
Энергетический обмен (диссимиляция — от лат. dissimilis ‒ несходный) — обратная ассимиляции сторона обмена веществ, совокупность реакций, которые приводят к высвобождению энергии химических связей. Это реакции расщепления жиров, белков, углеводов, нуклеиновых кислот до простых веществ.
Возможно три этапа диссимиляции: подготовительный, анаэробный и аэробный. Среда обитания определяет количество этапов диссимиляции. Их может быть три, если организм обитает в кислородной среде, и два, если речь идет об организме, обитающем в бескислородной среде (к примеру, в кишечнике).
Обсудим этапы энергетического обмена более подробно:
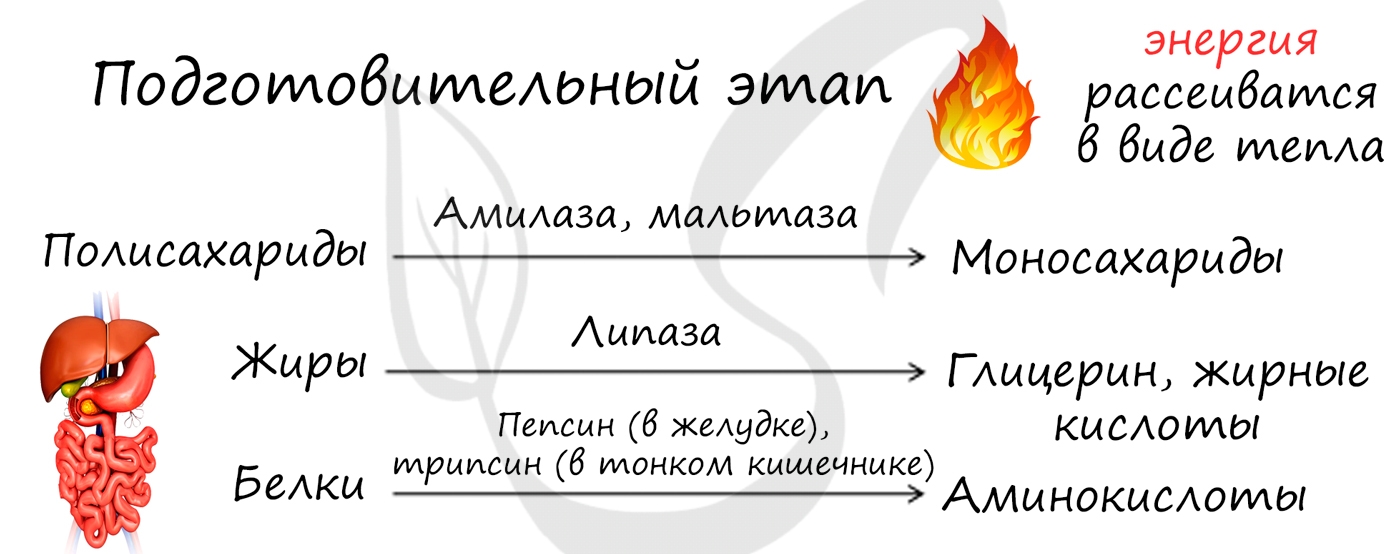
- Подготовительный этап
Подготовительный этап осуществляется ферментами в ЖКТ. В результате действия ферментов сложные вещества превращаются в более простые: полимеры распадаются на мономеры. Это сопровождается разрывом химических связей и выделением энергии, большая часть которой рассеивается в виде тепла.
Под действием ферментов белки расщепляются на аминокислоты, жиры — на глицерин и жирные кислоты, сложные углеводы — до простых сахаров.
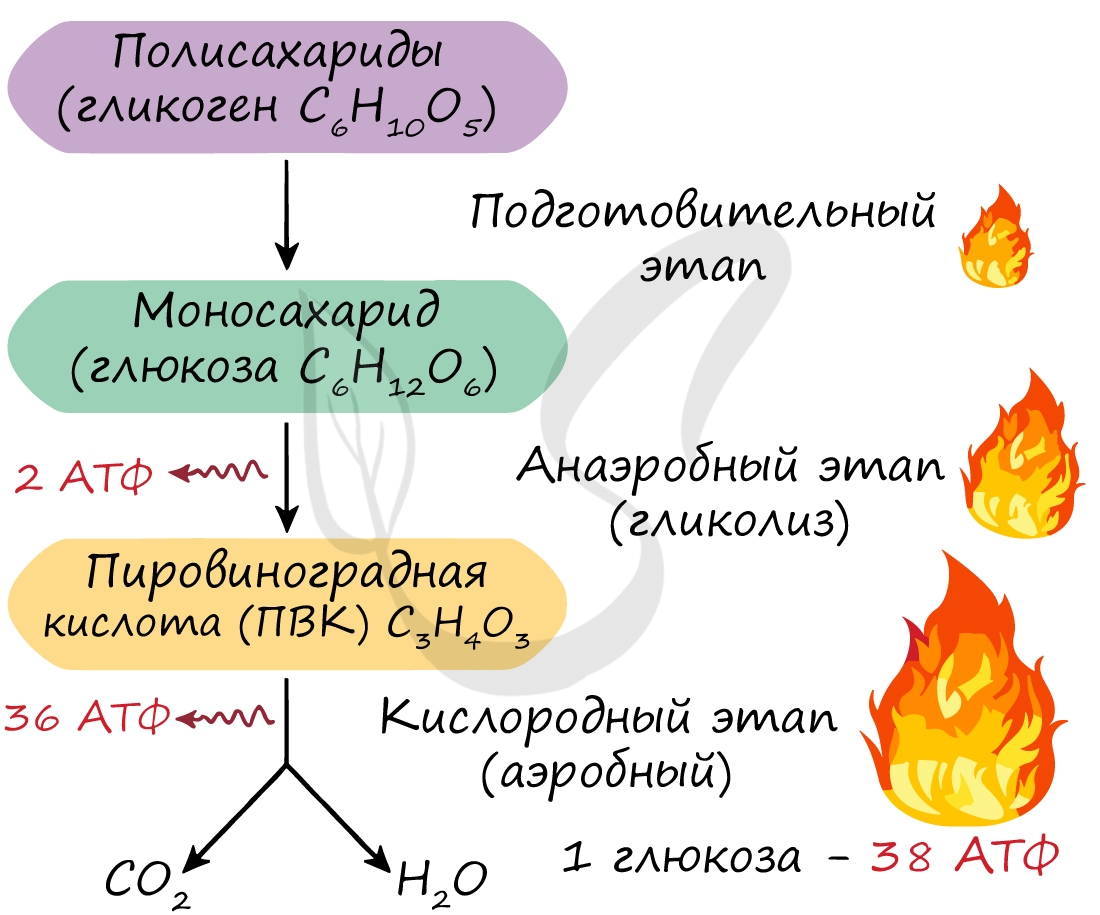
Этот этап является последним для организмов-анаэробов, обитающих в условиях, где кислород отсутствует. На этапе гликолиза происходит расщепление молекулы глюкозы: образуется 2 молекулы АТФ и 2 молекулы пировиноградной кислоты (ПВК). Происходит данный этап в цитоплазме клеток.
Кислородный этап (аэробный)
Этот этап доступен только для аэробов — организмов, живущих в кислородной среде. Из каждой молекулы ПВК, образовавшейся на этапе гликолиза, синтезируется 18 молекул АТФ — в сумме с двух ПВК выход составляет 36 молекул АТФ.
Таким образом, суммарно с одной молекулы глюкозы можно получить 38 АТФ (гликолиз + кислородный этап).
Кислородный этап протекает на кристах митохондрий (складках, выпячиваниях внутренней мембраны), где наибольшая концентрация окислительных ферментов. Главную роль в этом процессе играет так называемый цикл Кребса, который подробно изучает биохимия.
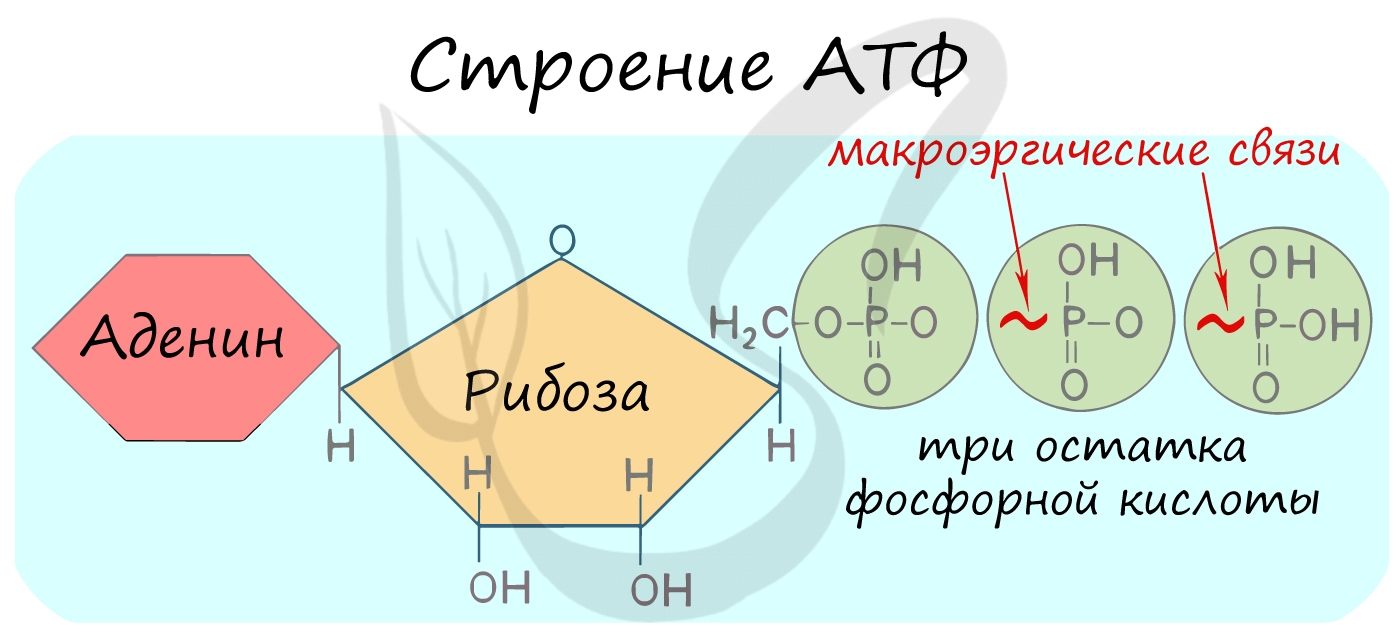
АТФ — аденозинтрифосфорная кислота
Трудно переоценить роль в клетке АТФ — универсального источника энергии. Молекула АТФ состоит из азотистого основания — аденина, углевода — рибозы и трех остатков фосфорной кислоты.
Между остатками фосфорной кислоты находятся макроэргические связи — ковалентные связи, которые гидролизуются с выделением большого количества энергии. Их принято обозначать типографическим знаком тильда «∽».
АТФ гидролизуется до АДФ (аденозиндифосфорная кислота), а затем и до АМФ (аденозинмонофосфорная кислота). Гидролиз АТФ сопровождается выделением энергии (E) на каждом этапе и может быть представлен такой схемой:
- АТФ + H2O = АДФ + H3PO4 + E
- АДФ + H2O = АМФ + H3PO4 + E
- АМФ + H2O = аденин + рибоза + H3PO4 + E
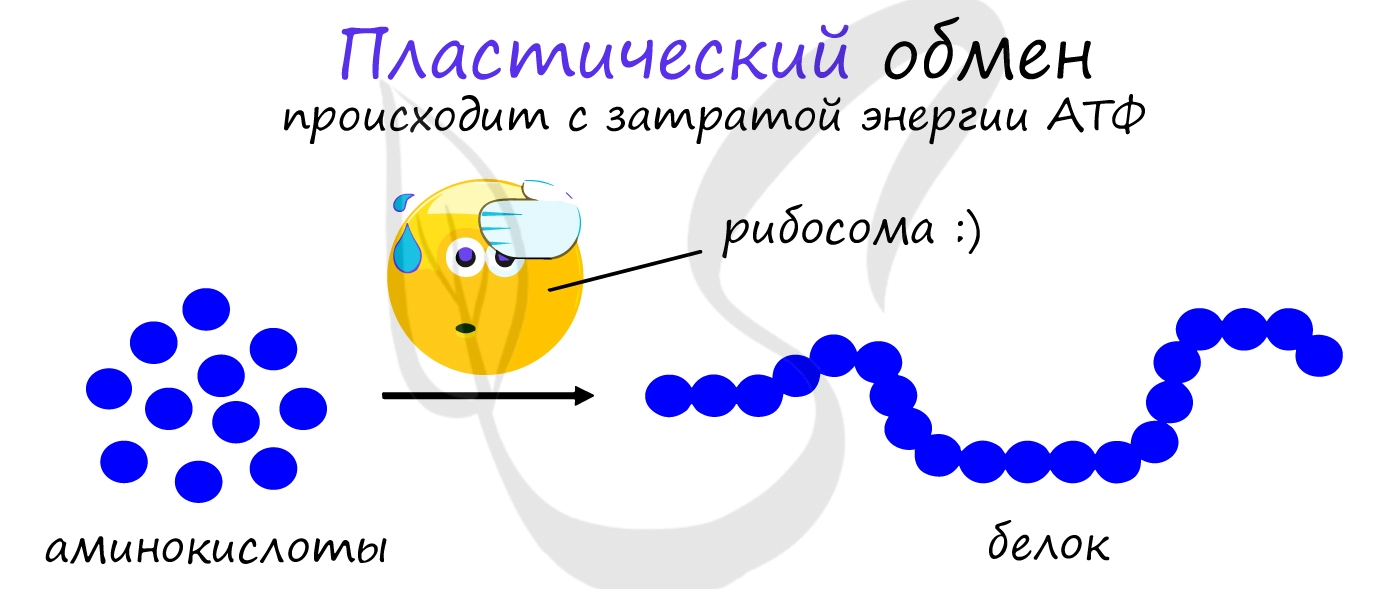
Пластический обмен
АТФ является универсальным источником энергии в клетке: энергия макроэргических связей АТФ используется для реакций пластического обмена (ассимиляции), протекающих с затратой энергии: синтеза белка на рибосоме (трансляции), удвоению ДНК (репликации) и т.д.
В результате пластического обмена в нашем организме происходит синтез белков, жиров и углеводов.
© Беллевич Юрий Сергеевич 2018-2022
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Метаболизм клетки. Энергетический обмен и фотосинтез. Реакции матричного синтеза.
Понятие метаболизма
Метаболизм — совокупность всех химических реакций, протекающих в живом организме. Значение метаболизма состоит в создании необходимых организму веществ и обеспечении его энергией.
Выделяют две составные части метаболизма — катаболизм и анаболизм.
Составные части метаболизма
Процессы пластического и энергетического обмена неразрывно связаны между собой. Все синтетические (анаболические) процессы нуждаются в энергии, поставляемой в ходе реакций диссимиляции. Сами же реакции расщепления (катаболизма) протекают лишь при участии ферментов, синтезируемых в процессе ассимиляции.
Роль ФТФ в метаболизме
Энергия, высвобождающаяся при распаде органических веществ, не сразу используется клеткой, а запасается в форме высокоэнергетических соединений, как правило, в форме аденозинтрифосфата (АТФ). По своей химической природе АТФ относится к мононуклеотидам.
АТФ (аденозинтрифосфорная кислота) — мононуклеотид, состоящий из аденина, рибозы и трёх остатков фосфорной кислоты, соединяющихся между собой макроэргическими связями.
В этих связях запасена энергия, которая высвобождается при их разрыве:
АТФ + H2O → АДФ + H3PO4 + Q1
АДФ + H2O → АМФ + H3PO4 + Q2
АМФ + H2O → аденин + рибоза + H3PO4 + Q3,
где АТФ — аденозинтрифосфорная кислота; АДФ — аденозиндифосфорная кислота; АМФ — аденозинмонофосфорная кислота; Q1 = Q2 = 30,6 кДж; Q3 = 13,8 кДж.
Запас АТФ в клетке ограничен и пополняется благодаря процессу фосфорилирования. Фосфорилирование — присоединение остатка фосфорной кислоты к АДФ (АДФ + Ф → АТФ). Он происходит с разной интенсивностью при дыхании, брожении и фотосинтезе. АТФ обновляется чрезвычайно быстро (у человека продолжительность жизни одной молекулы АТФ менее 1 мин).
Энергия, накопленная в молекулах АТФ, используется организмом в анаболических реакциях (реакциях биосинтеза). Молекула АТФ является универсальным хранителем и переносчиком энергии для всех живых существ.
Энергетический обмен
Энергию, необходимую для жизнедеятельности, большинство организмов получают в результате процессов окисления органических веществ, то есть в результате катаболических реакций. Важнейшим соединением, выступающим в роли топлива, является глюкоза.
По отношению к свободному кислороду организмы делятся на три группы.
Классификация организмов по отношению к свободному кислороду
| Группа | Характеристика | Организмы |
| Аэробы (облигатные аэробы) | Организмы, способные жить только в кислородной среде | Животные, растения, некоторые бактерии и грибы |
| Анаэробы (облигатные анаэробы) | Организмы, неспособные жить в кислородной среде | Некоторые бактерии |
| Факультативные формы (факультативные анаэробы) | Организмы, способные жить как в присутствии кислорода, так и без него | Некоторые бактерии и грибы |
У облигатных аэробов и факультативных анаэробов в присутствии кислорода катаболизм протекает в три этапа: подготовительный, бес- кислородный и кислородный. В результате органические вещества распадаются до неорганических соединений. У облигатных анаэробов и факультативных анаэробов при недостатке кислорода катаболизм протекает в два первых этапа: подготовительный и бескислородный. В результате образуются промежуточные органические соединения, еще богатые энергией.
Этапы катаболизма
1. Первый этап — подготовительный — заключается в ферментативном расщеплении сложных органических соединений на более простые. Белки расщепляются до аминокислот, жиры — до глицерина и жирных кислот, полисахариды — до моносахаридов, нуклеиновые кислоты — до нуклеотидов. У многоклеточных организмов это происходит в желудочно-кишечном тракте, у одноклеточных — в лизосомах под действием гидролитических ферментов. Высвобождающаяся при этом энергия рассеивается в виде теплоты. Образовавшиеся органические соединения либо подвергаются дальнейшему окислению, либо используются клеткой для синтеза собственных органических соединений.
2. Второй этап — неполное окисление (бескислородный) — заключается в дальнейшем расщеплении органических веществ, осуществляется в цитоплазме клетки без участия кислорода. Главным источником энергии в клетке является глюкоза. Бескислородное, неполное окисление глюкозы называется гликолизом. В результате гликолиза одной молекулы глюкозы образуется по две молекулы пировиноградной кислоты (ПВК, пируват) CH3COCOOH, АТФ и воды, а также атомы водорода, которые связываются молекулой-переносчиком НАД + и запасаются в виде НАД·Н.
Суммарная формула гликолиза имеет следующий вид:
C6H12O6 + 2H3PO4 + 2АДФ + 2НАД+ → 2C3Н4O3 + 2H2O + 2АТФ + 2НАД·Н.
Далее при отсутствии в среде кислорода продукты гликолиза (ПВК и НАД·Н) перерабатываются либо в этиловый спирт — спиртовое брожение (в клетках дрожжей и растений при недостатке кислорода)
CH3COCOOH → СО2 + СН3СОН
СН3СОН + 2НАД·Н → С2Н5ОН + 2НАД + ,
либо в молочную кислоту — молочнокислое брожение (в клетках животных при недостатке кислорода)
CH3COCOOH + 2НАД·Н → C3Н6O3 + 2НАД + .
При наличии в среде кислорода продукты гликолиза претерпевают дальнейшее расщепление до конечных продуктов.
3. Третий этап — полное окисление (дыхание) — заключается в окислении ПВК до углекислого газа и воды, осуществляется в митохондриях при обязательном участии кислорода.
Он состоит из трёх стадий:
А) образование ацетилкоэнзима А;
Б) окисление ацетилкоэнзима А в цикле Кребса;
В) окислительное фосфорилирование в электронотранспортной цепи.
А. На первой стадии ПВК переносится из цитоплазмы в митохондрии, где взаимодействует с ферментами матрикса и образует 1) диоксид углерода, который выводится из клетки; 2) атомы водорода, которые молекулами-переносчиками доставляются к внутренней мембране митохондрии; 3) ацетилкофермент А (ацетил-КоА).
Б. На второй стадии происходит окисление ацетилкоэнзима А в цикле Кребса. Цикл Кребса (цикл трикарбоновых кислот, цикл лимонной кислоты) — это цепь последовательных реакций, в ходе которых из одной молекулы ацетил-КоА образуются 1) две молекулы диоксида углерода, 2) молекула АТФ и 3) четыре пары атомов водорода, передаваемые на молекулы-переносчики — НАД и ФАД. Таким образом, в результате гликолиза и цикла Кребса молекула глюкозы расщепляется до СО2, а высвободившаяся при этом энергия расходуется на синтез 4 АТФ и накапливается в 10 НАД·Н и 4 ФАД·Н2.
В. На третьей стадии атомы водорода с НАД·Н и ФАД·Н2 окисляются молекулярным кислородом О2 с образованием воды. Один НАД·Н способен образовывать 3 АТФ, а один ФАД·Н2–2 АТФ. Таким образом, выделяющаяся при этом энергия запасается в виде ещё 34 АТФ.
Этот процесс протекает следующим образом. Атомы водорода концентрируются около наружной стороны внутренней мембраны митохондрии. Они теряют электроны, которые по цепи молекул-переносчиков (цитохромов) электронотранспортной цепи (ЭТЦ) переносятся на внутреннюю сторону внутренней мембраны, где соединяются с молекулами кислорода:
О2 + е — → О2 — .
В результате деятельности ферментов цепи переноса электронов внутренняя мембрана митохондрий изнутри заряжается отрицательно (за счёт О2 — ), а снаружи — положительно (за счёт Н + ), так что между её поверхностями создаётся разность потенциалов. Во внутреннюю мембрану митохондрий встроены молекулы фермента АТФ- синтетазы, обладающие ионным каналом. Когда разность потенциалов на мембране достигает критического уровня, положительно заряженные частицы H + силой электрического поля начинают проталкиваться через канал АТФазы и, оказавшись на внутренней поверхности мембраны, взаимодействуют с кислородом, образуя воду:
1/2О2 — +2H + → Н2О.
Энергия ионов водорода H + , транспортирующихся через ионный канал внутренней мембраны митохондрии, используется для фосфорилирования АДФ в АТФ:
АДФ + Ф → АТФ.
Такое образование АТФ в митохондриях при участии кислорода называется окислительным фосфорилированием.
Суммарное уравнение расщепления глюкозы в процессе клеточного дыхания:
C6H12O6 + 6O2 + 38H3PO4 + 38АДФ → 6CO2 + 44H2O + 38АТФ.
Таким образом, в ходе гликолиза образуются 2 молекулы АТФ, в ходе клеточного дыхания — ещё 36 молекул АТФ, в целом при пол- ном окислении глюкозы — 38 молекул АТФ.
Пластический обмен
Пластический обмен, или ассимиляция, представляет собой совокупность реакций, обеспечивающих синтез сложных органических соединений из более простых (фотосинтез, хемосинтез, биосинтез белка и др.).
Гетеротрофные организмы строят собственные органические вещества из органических компонентов пищи. Гетеротрофная ассимиляция сводится, по существу, к перестройке молекул:
органические вещества пищи (белки, жиры, углеводы) → простые органические молекулы (аминокислоты, жирные кислоты, моносахариды) → макромолекулы тела (белки, жиры, углеводы).
Автотрофные организмы способны полностью самостоятельно синтезировать органические вещества из неорганических молекул, потребляемых из внешней среды. В процессе фото- и хемосинтеза происходит образование простых органических соединений, из которых в дальнейшем синтезируются макромолекулы:
неорганические вещества (СО2, Н2О) → простые органические молекулы (аминокислоты, жирные кислоты, моносахариды) → макромолекулы тела (белки, жиры, углеводы).
Фотосинтез
Фотосинтез — синтез органических соединений из неорганических за счёт энергии света. Суммарное уравнение фотосинтеза:
Фотосинтез протекает при участии фотосинтезирующих пигментов, обладающих уникальным свойством преобразования энергии солнечного света в энергию химической связи в виде АТФ. Фотосинтезирующие пигменты представляют собой белковоподобные вещества. Наиболее важным является пигмент хлорофилл. У эукариот фотосинтезирующие пигменты встроены во внутреннюю мембрану пластид, у прокариот — во впячивания цитоплазматической мембраны.
Строение хлоропласта очень похоже на строение митохондрии. Во внутренней мембране тилакоидов гран содержатся фотосинтетические пигменты, а также белки цепи переноса электронов и молекулы фермента АТФ-синтетазы.
Процесс фотосинтеза состоит из двух фаз: световой и темновой.
1. Световая фаза фотосинтеза протекает только на свету в мембране тилакоидов граны.
К ней относятся поглощение хлорофиллом квантов света, образование молекулы АТФ и фотолиз воды.
Под действием кванта света (hv) хлорофилл теряет электроны, переходя в возбуждённое состояние:
Эти электроны передаются переносчиками на наружную, то есть обращенную к матриксу поверхность мембраны тилакоидов, где накапливаются.
Одновременно внутри тилакоидов происходит фотолиз воды, то есть её разложение под действием света:
Образующиеся электроны передаются переносчиками к молекулам хлорофилла и восстанавливают их. Молекулы хлорофилла возвращаются в стабильное состояние.
Протоны водорода, образовавшиеся при фотолизе воды, накапливаются внутри тилакоида, создавая Н + -резервуар. В результате внутренняя поверхность мембраны тилакоида заряжается положительно (за счёт Н + ), а наружная — отрицательно (за счёт е — ). По мере накопления по обе стороны мембраны противоположно заряженных частиц нарастает разность потенциалов. При достижении критической величины разности потенциалов сила электрического поля начинает проталкивать протоны через канал АТФ-синтетазы. Выделяющаяся при этом энергия используется для фосфорилирования молекул АДФ:
АДФ + Ф → АТФ.
Образование АТФ в процессе фотосинтеза под действием энергии света называется фотофосфорилированием.
Ионы водорода, оказавшись на наружной поверхности мембраны тилакоида, встречаются там с электронами и образуют атомарный водород, который связывается с молекулой-переносчиком водорода НАДФ (никотинамидадениндинуклеотидфосфат):
2Н + + 4е – + НАДФ + → НАДФ·Н2.
Таким образом, во время световой фазы фотосинтеза происходят три процесса: образование кислорода вследствие разложения воды, синтез АТФ и образование атомов водорода в форме НАДФ·Н2. Кислород диффундирует в атмосферу, а АТФ и НАДФ·Н2 участвуют в процессах темновой фазы.
2. Темновая фаза фотосинтеза протекает в матриксе хлоропласта как на свету, так и в темноте и представляет собой ряд последовательных преобразований СО2, поступающего из воздуха, в цикле Кальвина. Осуществляются реакции темновой фазы за счёт энергии АТФ. В цикле Кальвина СО2 связывается с водородом из НАДФ·Н2 с образованием глюкозы.
В процессе фотосинтеза кроме моносахаридов (глюкоза и др.) синтезируются мономеры других органических соединений — аминокислоты, глицерин и жирные кислоты. Таким образом, благодаря фотосинтезу растения обеспечивают себя и всё живое на Земле необходимыми органическими веществами и кислородом.
Сравнительная характеристика фотосинтеза и дыхания эукариот представлена в таблице.
Сравнительная характеристика фотосинтеза и дыхания эукариот
Генетическая информация у всех организмов хранится в виде определённой последовательности нуклеотидов ДНК (или РНК у РНК-содержащих вирусов). Прокариоты содержат генетическую информацию в виде одной молекулы ДНК. В эукариотических клетках генетический материал распределён в нескольких молекулах ДНК, организованных в хромосомы.
ДНК состоит из кодирующих и некодирующих участков. Кодирующие участки кодируют РНК. Некодирующие области ДНК выполняют структурную функцию, позволяя участкам генетического материала упаковываться определённым образом, или регуляторную функцию, участвуя во включении генов, направляющих синтез белка.
Кодирующими участками ДНК являются гены. Ген — участок молекулы ДНК, кодирующей синтез одной мРНК (и соответственно полипептида), рРНК или тРНК.
Участок хромосомы, где расположен ген называется локусом. Совокупность генов клеточного ядра представляет собой генотип, совокупность генов гаплоидного набора хромосом — гено́м, совокупность генов внеядерных ДНК (митохондрий, пластид, цитоплазмы) — плазмон.
Реализация информации, записанной в генах, через синтез белков называется экспрессией (проявлением) генов. Генетическая информация хранится в виде определённой последовательности нуклеотидов ДНК, а реализуется в виде последовательности аминокислот в белке. Посредниками, переносчиками информации выступают РНК. То есть реализация генетической информации происходит следующим образом:
ДНК → РНК → белок.
Этот процесс осуществляется в два этапа:
1) транскрипция;
2) трансляция.
Транскрипция (от лат. transcriptio — переписывание) — синтез РНК с использованием ДНК в качестве матрицы. В результате образуются мРНК, тРНК и рРНК. Процесс транскрипции требует больших затрат энергии в виде АТФ и осуществляется ферментом РНК-полимеразой.
Одновременно транскрибируется не вся молекула ДНК, а лишь отдельные её отрезки. Такой отрезок (транскриптон) начинается промотором — участком ДНК, куда присоединяется РНК-полимераза и откуда начинается транскрипция, а заканчивается терминатором — участком ДНК, содержащим сигнал окончания транскрипции. Транскриптон — это ген с точки зрения молекулярной биологии.
Транскрипция, как и репликация, основана на способности азотистых оснований нуклеотидов к комплементарному связыванию. На время транскрипции двойная цепь ДНК разрывается, и синтез РНК осуществляется по одной цепи ДНК.
В процессе транскрипции последовательность нуклеотидов ДНК переписывается на синтезирующуюся молекулу мРНК, которая выступает в качестве матрицы в процессе биосинтеза белка.
Гены прокариот состоят только из кодирующих нуклеотидных последовательностей.
Гены эукариот состоят из чередующихся кодирующих (экзонов) и некодирующих (интронов) участков.
После транскрипции участки мРНК, соответствующие интронам, удаляются в ходе сплайсинга, являющегося составной частью процессинга.
Процессинг — процесс формирования зрелой мРНК из её предшественника пре-мРНК. Он включает два основных события. 1.Присоединение к концам мРНК коротких последовательностей нуклеотидов, обозначающих место начала и место конца трансляции. Сплайсинг — удаление неинформативных последовательностей мРНК, соответствующих интронам ДНК. В результате сплайсинга молекулярная масса мРНК уменьшается в 10 раз. Трансляция (от лат. translatio — перевод) — синтез полипептидной цепи с использованием мРНК в роли матрицы.
В трансляции участвуют все три типа РНК: мРНК является информационной матрицей; тРНК доставляют аминокислоты и узнают кодоны; рРНК вместе с белками образуют рибосомы, которые удерживают мРНК, тРНК и белок и осуществляют синтез полипептидной цепи.
Этапы трансляции
Реакции матричного синтеза. К реакциям матричного синтеза относятся
- самоудвоение ДНК (репликация);
- образование мРНК, тРНК и рРНК на молекуле ДНК (транскрипция);
- биосинтез белка на мРНК (трансляция).
Все эти реакции объединяет то, что молекула ДНК в одном случае или молекула мРНК в другом выступают в роли матрицы, на которой происходит образование одинаковых молекул. Реакции матричного синтеза являются основой способности живых организмов к воспроизведению себе подобных.
Регуляция экспрессии генов. Тело многоклеточного организма построено из разнообразных клеточных типов. Они отличаются структурой и функциями, то есть дифференцированы. Различия проявляются в том, что помимо белков, необходимых любой клетке организма, клетки каждого типа синтезируют ещё и специализированные белки: в эпидермисе образуется кератин, в эритроцитах — гемоглобин и т. д. Клеточная дифференцировка обусловлена изменением набора экспрессируемых генов и не сопровождается какими-либо необратимыми изменениями в структуре самих последовательностей ДНК.
Уравнения энергетического обмена для егэ

Видео YouTube

СТАДИИ ЭНЕРГЕТИЧЕСКОГО ОБМЕНА
Энергетический обмен (катаболизм, диссимиляция) — это процессы расщепления ве ществ с высвобождением энергии. Высвобожденная энергия преобразуется в энергию АТФ. Наиболее важными процессами энергетического обмена являются дыхание и брожение.

Энергетический обмен – это совокупность химических реакций постепенного распада органических соединений, сопровождающихся высвобождением энергии, часть которой расходуется на синтез АТФ. Синтезированная АТФ становится универсальным источником энергии для жизнедеятельности организмов. Она образуется в результате реакции фосфорилирования – присоединения остатков фосфорной кислоты к молекуле АДФ. На эту реакцию расходуется энергия, которая затем накапливается в макроэргических связях молекулы АТФ, при распаде молекулы АТФ или при ее гидролизе до АДФ клетка получает около 40 кДж энергии.
АТФ – постоянный источник энергии для клетки, она мобильно может доставлять химическую энергию в любую часть клетки. Когда клетке необходима энергия – достаточно гидролизовать молекулу АТФ. Энергия выделяется в результате реакции диссимиляции (расщепления органических веществ), в зависимости от специфики организма и условий его обитания энергетический обмен проходит в два или три этапа. Большинство живых организмов относятся к аэробам, использующим для обмена веществ кислород, который поступает из окружающей среды. Для аэробов энергетический обмен проходит в три этапа:
В организмах, которые обитают в бескислородной среде и не нуждаются в кислороде для энергетического обмена – анаэробах и аэробах, при недостатке кислорода проходят энергетический обмен в два этапа:
Количество энергии, которое выделяется при двухэтапном варианте намного меньше, чем в трехэтапном.
ЭТАПЫ ЭНЕРГЕТИЧЕСКОГО ОБМЕНА
Подготовительный этап – во время него крупные пищевые полимерные молекулы распадаются на более мелкие фрагменты. В желудочно-кишечном тракте многоклеточных организмов он осуществляется пищеварительными ферментами, у одноклеточных – ферментами лизосом. Полисахариды распадаются на ди- и моносахариды, белки – до аминокислот, жиры – до глицерина и жирных кислот. В ходе этих превращений энергии выделяется мало, она рассеивается в виде тепла, и АТФ не образуется. Образующиеся в ходе подготовительного этапа соединения-мономеры могут участвовать в реакциях пластического обмена (в дальнейшем из них синтезируются вещества, необходимые для клетки) или подвергаться дальнейшему расщеплению с целью получения энергии.
Большинство клеток в первую очередь используют углеводы, жиры остаются в первом резерве и используются по окончания запаса углеводов. Хотя есть и исключения: в клетках скелетных мышц при наличии жирных кислот и глюкозы предпочтение отдается жирным кислотам. Белки расходуются в последнюю очередь, когда запас углеводов и жиров будет исчерпан – при длительном голодании.
Бескислородный этап (гликолиз) – происходит в цитоплазме клеток. Главным источником энергии в клетке является глюкоза. Ее бескислородное расщепление называют анаэробным гликолизом. Он состоит из ряда последовательных реакций по превращению глюкозы в лактат. Его присутствие в мышцах хорошо известно уставшим спортсменам. Этот этап заключается в ферментативном расщеплении органических веществ, полученных в ходе первого этапа. Так как глюкоза является наиболее доступным субстратом для клетки как продукт расщепления полисахаридов, то второй этап можно рассмотреть на примере ее бескислородного расщепления – гликолиза (Рис. 1).

Рис. 1. Бескислородный этап
Гликолиз – многоступенчатый процесс бескислородного расщепления молекулы глюкозы, содержащей шесть атомов углерода, до двух молекул пировиноградной кислоты (пируват). Реакция гликолиза катализируется многими ферментами и протекает в цитоплазме клетки. В ходе гликолиза при расщеплении одного моля глюкозы выделяется около 200 кДж энергии, 60 % ее рассеивается в виде тепла, 40 % – для синтезирования двух молекул АТФ из двух молекул АДФ. При наличии кислорода в среде пировиноградная кислота из цитоплазмы переходит в митохондрии и участвует в третьем этапе энергетического обмена. Если кислорода в клетке нет, то пировиноградная кислота преобразуется в животных клетках или превращается в молочную кислоту.
В микроорганизмах, которые существуют без доступа кислорода – получают энергию в процессе брожения, начальный этап аналогичен гликолизу: распад глюкозы до двух молекул пировиноградной кислоты, и далее она зависит от ферментов, которые находятся в клетке – пировиноградная кислота может преобразовываться в спирт, уксусную кислоту, пропионовую и молочную кислоту. В отличие от того, что происходит в животных тканях, у микроорганизмов этот процесс носит название молочнокислого брожения. Все продукты брожения широко используются в практической деятельности человека: это вино, квас, пиво, спирт, кисломолочные продукты. При брожении, так же как и при гликолизе, выделяется всего две молекулы АТФ.
Кислородный этап стал возможен после накопления в атмосфере достаточного количества молекулярного кислорода, он происходит в митохондриях клеток. Он очень сложен по сравнению с гликолизом, это процесс многостадийный и идет при участии большого количества ферментов. В результате третьего этапа энергетического обмена из двух молекул пировиноградной кислоты формируется углекислый газ, вода и 36 молекул АТФ (Рис. 2).

Рис. 2. Митохондрия
Две молекулы АТФ запасаются в ходе бескислородного расщепления молекулами глюкозы, поэтому суммарный энергетический обмен в клетке в случае распада глюкозы можно представить как:
С 6 Н 12 О 6 + 6О 2 + 38АДФ + 38Н 3 РО 4 = 6СО 2 + 44Н 2 О + 38АТФ
В результате окисления одной молекулы глюкозы шестью молекулами кислорода образуется шесть молекул углекислого газа и выделяется тридцать восемь молекул АТФ.
Мы видим, что в трехэтапном варианте энергетического обмена выделяется гораздо больше энергии, чем в двухэтапном варианте – 38 молекул АТФ против 2.



В отсутствие кислорода или при его недостатке про исходит брожение. Брожение является эволюционно бо лее ранним способом получения энергии, чем дыхание, однако оно энергетически менее выгодно, поскольку ко нечными продуктами брожения являются органические вещества, богатые энергией. Существует несколько видов брожения, названия которых определяются конечными продуктами: молочнокислое, спиртовое, уксуснокислое и др. Так, в скелетных мышцах в отсутствие кислорода протекает молочнокислое брожение, в ходе которого пировиноградная кислота восстанавли вается до молочной кислоты. При этом восстановленные ранее коферменты НАДН расходу ются на восстановление пирувата:

Для многих микроорганизмов брожение является единственным способом ассимиля ции энергии. Большинство таких организмов живет в анаэробных условиях и погибает в присутствии кислорода, но есть и такие, которые нормально существуют и в присутствии кислорода, и без него. Например, дрожжевые грибы при спиртовом брожении окисляют пировиноградную кислоту до этилового спирта и оксида углерода (IV):
http://examer.ru/ege_po_biologii/teoriya/metabolizm_kletki
http://www.sites.google.com/site/biologiaege/obmen-vesestv/energeticeskij-obmen